
Cualquiera a quien su disposición le conduzca a atribuir más peso a las dificultades no explicadas que a determinado número de hechos, rechazará sin duda mi teoría.
CHARLES DARWIN
El origen de las especies, 1859
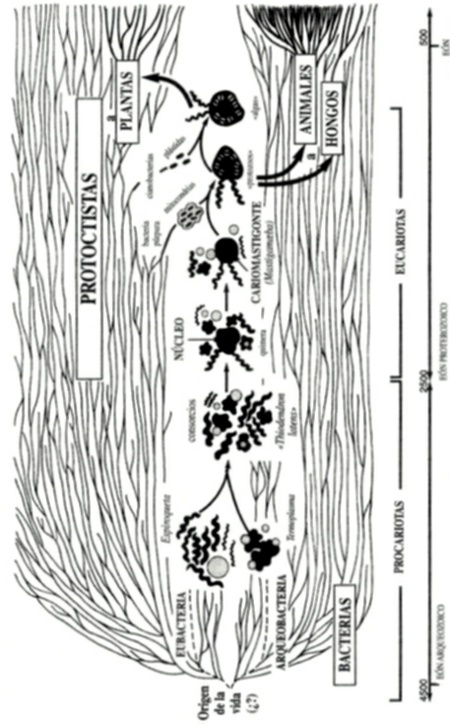
Figura FM. 1 Origen bacteriano de las células nucleadas
Los autores de Captando genomas contravienen esta tendencia, demostrando la descomunal importancia de las interacciones entre individuos de especies distintas. Gran parte del progreso de la evolución se debe al establecimiento de consorcios entre dos organismos con genomas completamente diferentes. Los ecólogos han comenzado apenas a describir estas interacciones.
Entre los millones de interacciones posibles —incluyendo el parasitismo—, los autores han seleccionado una como objeto principal de su trabajo: la simbiosis. Éste es el nombre que recibe la interacción mutua que involucra la asociación física entre «organismos denominados de forma diferente». Los líquenes constituyen el ejemplo clásico de simbiosis. En ellos, un hongo se asocia con un alga o con una cianobacteria. Considerada inicialmente la simbiosis como algo excepcional, con el tiempo se descubrió su naturaleza prácticamente universal. Por ejemplo, los microbios que viven en una de las cavidades del estómago de la vaca, proporcionando las enzimas necesarias para la digestión de la celulosa, son simbiontes de la vaca. Desde hace tiempo, Lynn Margulis se encuentra en la vanguardia del estudio sobre la simbiosis. Consiguió convencer a los citólogos de que las mitocondrias son simbiontes de las células, tanto vegetales como animales, del mismo modo que los cloroplastos lo son de las primeras. El surgimiento de una nueva forma a partir de esta clase de simbiosis es conocida bajo el nombre de simbiogénesis.
Durante muchos años Margulis ha liderado la interpretación de entidades evolutivas como productos de la simbiogénesis. El acontecimiento de esta índole más asombroso —y para muchos aún increíble— consiste en el origen de las eucariotas[1] mediante la fusión de una arqueobacteria con alguna eubacteria. Ambos progenitores contribuyeron al resultado con importantes capacidades fisiológicas, lo que garantizaría el extraordinario éxito evolutivo de las células eucariotas, que constituyen los elementos de construcción de la totalidad de animales, plantas y hongos.
La simbiogénesis constituye el tema central de la presente obra. Los autores demuestran en ella, de forma convincente, que una proporción sorprendentemente elevada de linajes evolutivos tuvo sus orígenes en la simbiogénesis. En esos casos, la combinación de dos genomas totalmente distintos forma un consorcio que se convierte en un objetivo de selección como entidad única. Por la estabilidad mutua de la relación de este modo establecida, la simbiosis difiere de otras clases de interacción como el carnivorismo, el herbivorismo y el parasitismo.
La adquisición de un nuevo genoma puede ser tan instantánea como el acontecimiento cromosomático que conduce a la poliploidía [2]. Los autores nos incitan a la sugerencia de que esta clase de acontecimiento pudiera contradecir el principio darwiniano de evolución gradual. En realidad, la incorporación de un nuevo genoma puede muy bien constituir un proceso muy lento, que se extienda a lo largo de numerosas generaciones. Pero aun en el caso de que fuera instantáneo, nunca sería más brusco que cualquier otro suceso conducente a la poliploidía.
Los autores se refieren al acto de la simbiogénesis como ejemplo de especiación. Algunas de sus afirmaciones tal vez puedan conducir al lector poco informado a la conclusión errónea de que la especiación se debe en todos los casos a la simbiogénesis. La realidad es otra: la especiación —la multiplicación de especies— y la simbiogénesis son dos procesos independientes y sobrepuestos. No existe indicio alguno de que ninguna de las 10.000 especies de aves o de las 4.500 especies de mamíferos se hayan originado por medio de la simbiogénesis.
Otra de las interpretaciones evolutivas contenidas en la presente obra es igualmente vulnerable. Sus autores sugieren que la incorporación de nuevos genomas en casos de simbiogénesis restituye la validez del venerable principio de herencia de caracteres adquiridos, conocido como «herencia lamarquiana». Eso no es cierto. Ambos procesos son enteramente diferentes. La herencia lamarquiana consiste en la transmisión de fenotipos modificados, mientras que la simbiogénesis implica la transferencia de partes de genomas incorporadas.
Tal vez el mayor mérito de la presente obra resida en su capacidad para introducir al lector en el mundo de los microbios. No puede negarse que el estudioso medio de animales y plantas sabe más bien poco — ¡o casi nada!— de este mundo. Margulis y Sagan no limitan su trabajo a la simbiogénesis, sino que nos ofrecen asimismo una apasionante descripción de protistas y bacterias. ¿Qué biólogo es sabedor de que, mientras que los animales disponen de un conjunto único de itinerarios metabólico, diversas clases de bacterias tienen más de veinte a su disposición, todos ellos fundamentalmente diferentes entre sí? Mientras que los especialistas dedicados al estudio de la fisiología de los vertebrados se cuentan por millares, las fisiologías de las bacterias, mucho más diversas, constituyen el ámbito de estudio de tan sólo un puñado de microbiólogos.
Los capítulos 9, 10 y 11 están dedicados a fascinantes descripciones de docenas de consorcios inusuales de plantas y animales con microbios (protistas, hongos inferiores y bacterias). No deja de ser admirable el modo en que la selección natural ha tenido éxito en la fusión en consorcios de las combinaciones más improbables. Ninguna ficción podría resultar más fascinante que algunos de estos casos de simbiogénesis.
Habida cuenta de la dedicación de los autores a su campo específico, no resulta sorprendente que, en ocasiones, lleguen a interpretaciones que algunos podamos considerar discutibles. El lector sabrá pasar por alto aquellas que contradigan claramente los descubrimientos de la biología moderna. En cambio, podrá concentrarse en otras interpretaciones brillantes y novedosas contenidas en este texto, agradeciendo a sus autores que hayan llamado nuestra atención sobre manifestaciones de vida que, a pesar de su importancia en el conjunto del edificio de la Naturaleza, suelen ser sistemáticamente ignoradas por la mayoría de los biólogos.
No olvidemos pues nunca la importante lección que Margulis y Sagan nos transmiten: el mundo de la vida no consiste únicamente en especies independientes, sino que cada individuo de la mayoría de las especies es, en realidad, un consorcio de varias de ellas. Las relaciones entre organismos superiores y microbios son infinitas en número y, en la mayoría de los casos, constituyen una contribución indispensable para el bienestar de ambos progenitores. Algún conocimiento acerca de esta vasta rama de la biología debería ser parte ineludible de la educación de todo biólogo. La biología es mucho más que ratas, Drosophila, Caenorhabditis y Escherichia coli. El estudio de la simbiogénesis puede contribuir a una comprensión más profunda del universo de la vida. Para avanzar en su conocimiento, nada mejor que adentrarse en las páginas de Captando genomas.
ERNST MAYR
Bedford, Massachusetts
Febrero del 2002
Nada tienen que ver con ello ni ninguna deidad omnipotente ni ninguna fantástica cantidad de buena suerte. La historia que comienza con la aplicación de Jean Baptiste Lamarck, los datos recopilados por Charles Darwin, los guisantes de Gregor Mendel, el tesón de James Watson y la falta de humildad de Francis Crick termina, con una especie de ironía romántica, en el humus y en el lodo. Sus protagonistas son bacterias rápidas y decididas, junto con expertos arquitectos protistas sobre una Tierra tectónicamente activa bajo un Sol energético. Guerras, alianzas, extraños encuentros sexuales, uniones, treguas y victorias constituyen los dramas de esta historia. Mutaciones aleatorias de ADN, de consecuencias originalmente destructivas, dan cuenta tan sólo de sus inicios. El papel de la aleatoriedad ha sido exagerado a lo largo de la saga de la evolución. Fármacos, compuestos químicos y otras moléculas no son más que estimulantes y componentes. Los seres vivos, por el contrario, son sus verdaderos protagonistas.
Al igual que el disco duro del ordenador, la molécula de ADN almacena información evolutiva pero no la crea. Puesto que no son “entidades” en un sentido coherente, los genes egoístas pueden ser comparados a invenciones de una imaginación hiperactiva, inicialmente anglófona. El verdadero ser es la célula viva, entidad que no puede evitar crear copias de sí misma. Ella es el personaje central. El motor de la evolución se mueve gracias a seres diminutos de cuya existencia apenas somos conscientes. Les tememos y los maldecimos, pero las bacterias, estos seres vivos y diminutos, escapan a nuestra atención salvo en esas ocasiones temibles en que nos alarman o nos amenazan. Ignoramos o desdeñamos el hecho de que tienen vida propia. Pero la tienen. Las acciones de las bacterias y de otros seres invisibles perpetúan y generan especies viejas y nuevas.
Tanto si es sábado noche como si es cualquier otra, sucede algo espectacular. Este espectáculo ha estado en escena al menos durante 3.000 millones de años. Sus primeros actos son demasiado importantes como para que los ignoremos. Tan sólo muy al final de esta larga historia adquiere protagonismo ese simio social que anda erguido y que charla sin cesar. Ese simio desprovisto casi por completo de pelo, que corta selvas tropicales con soltura, se engaña con visiones de su propia importancia. Su sentido de unicidad y sus intereses egoístas, combinados con su prolijidad tecnológica, han conducido a una explosión demográfica de primates sin precedentes. Pero esta historia no trata de este simio, de sus cópulas, de sus convecinos ni de sus víctimas, sino más bien de sus antepasados no humanos. Mucho antes de las hogueras de campamento, de las proclamas de independencia, de las ciudades y de la expansión urbana, la Tierra giraba ya alrededor del Sol habitada por innumerables variedades de vida superficialmente distinta. La saga evolutiva entera sobre cómo las especies se originaron y se extinguieron puede constituir la narración más grande jamás contada. Después de todo, es la historia de todos y cada uno de nosotros.
Parte I
La evolución como imperativo
Capítulo 1
Darwinismo si, neodarwinismo, no
La evolución de DarwinIrónicamente, El origen de las especies —obra fundamental de Charles Darwin—, que presentaba a la comunidad científica y al gran público por igual pruebas abrumadoras de la teoría de la selección natural, no explica en ningún momento de dónde proceden las nuevas especies.
La simbiosis como fuente de variación hereditaria
El mito del individuo independiente
Las especies son denominaciones que se otorgan a organismos extremadamente parecidos, sean éstos animales, plantas, hongos o microorganismos. Habida cuenta de nuestra necesidad de identificar venenos, predadores, materiales útiles para construir cobijos, combustible, alimento y demás necesidades, desde antaño hemos venido adjudicando nombres a objetos vivos u otrora vivos. Los nombres de especies de organismos que probablemente conozcas están relacionados en la tabla 1.1. Hasta el Renacimiento, no obstante, los nombres de seres vivos variaban de lugar en lugar y raramente estaban definidos con precisión. Esta confusión generalizada de nombres locales y descripciones inconsistentes llevó al naturalista sueco Carolus von Linneo (1709-1789) a tratar de incorporar rigor y comprensibilidad internacional a esas descripciones. Desde que Linnaeus (su nombre en latín; Linneo en castellano) impusiera orden sobre unas 10.000 especies de seres vivos, los científicos utilizan un primer nombre (el género o grupo mayor, más inclusivo) seguido de un segundo nombre (el de la especie o grupo más reducido y menos inclusivo) para referirse a los organismos, sean éstos vivos o fósiles.

Los fósiles son restos muertos, pruebas de vida pasada. La palabra procede del francés fosse, que significa zanja, excavación [3]. Como sucede en el caso del enigmático trilobite Paradoxides paradoxismus, también lo fósiles reciben nombres y son agrupados en base a sus semejanzas y diferencias morfológicas.
Al igual que “espectáculo” o “especial”, la palabra “especie” procede del latín speculor (ver, mirar). A sabiendas o no, todos utilizamos el concepto morfológico de especie: los perros tienen aspecto de perro, son perros y están todos ellos clasificados como Canis familiaris. Los problemas surgen cuando tratamos de nombrar a los coyotes (Canis latrans), a los lobos (Canis lupus o lobo gris; Canis rufas o lobo rojo), o a otros animales estrechamente relacionados.
Los zoólogos, cuya profesión consiste en estudiar a animales, han impuesto un concepto distinto de especie, que denominan «concepto de especie biológica». En estado natural, coyotes y perros no se aparean para producir descendencia plenamente fértil, sino que se encuentran «reproductivamente aislados». La definición zoológica de las especies se refiere a organismos que pueden hibridar, es decir, que pueden aparearse y producir descendencia fértil. Así pues, los organismos que se aparean (personas, toros y vacas, etc.) pertenecen a la misma especie. A los botánicos, que se ocupan del estudio de las plantas, esta definición también les resulta útil.
Está actualmente en boga un tercer concepto de especie, el “filogenético”, “evolutivo” o “cladístico”. Según este concepto, los grupos de organismos (personas, plantas de maíz o gallinas) considerados como descendientes de unos mismos antepasados (“clades”) son clasificados como miembros de una misma especie. Estos organismos reciben la calificación de “monofílicos” por el hecho de descender de «un mismo y único antepasado».
Todos tenemos un fuerte sentido de la especie; para sobrevivir, nuestros antepasados necesitaban poder reconocer el alimento, la pareja potencial, la paja, las serpientes venenosas y muchos otros organismos. A los críos de dos años les encanta reconocer animales domésticos, pájaros e incluso peces, no hay más que ver el éxito de los ositos de peluche y los dinosaurios de juguete. Para la supervivencia, tanto de nuestra propia especie como de otras muchas, ha sido crucial cierta cognición evolutiva instintiva.
La evolución, el estudio de los cambios de la vida a lo largo del tiempo, consiste en gran medida en el seguimiento de las huellas de los orígenes de las especies. Argumentaremos aquí por qué creemos que, de estos tres conceptos de especie, el morfológico tradicional sigue siendo el más adecuado. Como demostraremos, la especie morfológica constituye la manifestación externa de la especie simbiogenética.
La asignación de animales similares de géneros compatibles a una misma especie, a condición de que puedan emparejarse y producir descendencia viable en el medio natural, resulta adecuada para mamíferos y muchos otros animales, pero no es general. Ampliaremos aquí el concepto de especiación para dar cabida en él a la totalidad de los organismos. He aquí nuestra definición simbiogenética de las especies: sugerimos que, si el organismo A es de la misma especie que el organismo B, ambos tienen que estar compuestos por los mismos conjuntos de genomas integrados, tanto cualitativa como cuantitativamente. Todos los organismos que puedan ser asignados a una única especie son productos de la simbiogénesis; en otras palabras, que A y B son asignados a la misma especie porque comparten la misma cantidad de las mismas diferentes clases de genomas integrados. Las bacterias carecen de especies porque ninguna de ellas (ni siquiera las eubacterias o las arqueobacterias) evolucionó a partir de la integración simbiótica de células anteriormente independientes. El proceso de especiación se inicia con las primeras eucariotas (los primeros protistas u organismos con núcleo). El concepto de que todas las bacterias son fértiles entre sí —es decir, que pueden transferirse genes de una a otra por muy distintos que sean los individuos recombinantes implicados en la transmisión— ha quedado ampliamente demostrado a lo largo de más de treinta años, siendo de nuevo explicado por Sonea y Matthieu en el 2000, en su libro Prokaryotology. Irónicamente, el popular punto de vista evolucionista consistente en que los organismos evolucionan por la acumulación de mutaciones aleatorias, resulta más adecuado para describir el proceso evolutivo de las bacterias. Todos los demás organismos superiores, más conocidos, se originaron por medio de integraciones simbióticas que dieron lugar a asociaciones permanentes. Los componentes simbióticos, previamente separados, se integraban biológicamente para originar individuos completamente nuevos, siempre en poblaciones. Como podemos comprobar en el Capítulo 12 con el trabajo de fisión cariotípica, los anteriores simbiontes quedaron tan desfigurados en su morfología y en sus capacidades, que su verdadera naturaleza tan sólo puede ser revelada tras una minuciosa investigación. Sea cual fuere el origen de la variación evolutiva objeto de estudio, es la selección natural la que incesantemente elimina los seres cuya forma, cuya fisiología, cuyo comportamiento y cuya química no resultan adecuados para un medio dado en un tiempo y en un lugar determinados, sean cuales fueren sus detalles. Las células bacterianas tienen un único genoma que adquirió sus conjuntos de genes —habitualmente mil o más— de procariotas, de uno en uno o de dos en dos. Los eucariotas adquieren e integran genomas completos para formar “individuos”. Por ejemplo, todas las células vegetales tienen como mínimo cuatro genomas integrados:
- las eubacterias móviles y
- las arqueo-bacterias sintetizadoras de proteínas que formaron la célula nucleada (el primer protista) seguidas de
- las protobacterias respiradoras de oxígeno que se convirtieron en el mitocondrio y
- las cianobacterias que se transformaron en el cloroplasto.
Sin embargo, el concepto de especie “filogenético”, “evolutivo” o “cladístico” está completamente mal enfocado, y su adopción interfiere con la capacidad para comprender el surgimiento de las especies. La simbiosis a largo plazo, que conduce al origen de las especies por simbiogénesis, requiere la integración de al menos dos organismos de denominaciones distintas. Ningún organismo o grupo de organismos visibles «desciende de un único antepasado común».
El propósito de esta obra consiste en explicar, mediante pruebas abundantes acumuladas por científicos de todo el mundo, este nuevo concepto sobre cómo las nuevas especies llegaron realmente a la existencia.
§. La evolución de Darwin
“Evolución” es un término que Charles Darwin nunca utilizó. En lugar de ello se refirió a «descendencia con modificaciones». Aún así, el concepto básico moderno de evolución es sin duda atribuible a él. Si levantamos la mano para jurar que somos darwinistas, estamos jurando también, con idéntico fervor, que no estamos de acuerdo con sus seguidores neodarwinistas ni con otros evolucionistas modernos. Es necesario distinguir los puntos de vista originales de Darwin de los de sus sucesores. Pero habida cuenta de que, tanto el neodarwinismo como la “síntesis moderna” que atribuye el cambio evolutivo a la mutación aleatoria, fueron desarrollados entre 1930 y 1960, mucho después de la muerte de Darwin en 1881, de ningún modo pudo él tener relación alguna con ambas posturas, del mismo modo que tampoco podía saber nada acerca de la “biología evolutiva”. La “síntesis moderna” fue la ciencia inventada para unificar la idea de Darwin acerca del cambio gradual de organismos y poblaciones, con el concepto de Gregor Mendel de estasis genética.
¿Cuáles son las diferencias entre el darwinismo y las ideas modernas predominantes? ¿En qué difieren los conceptos aquí presentados con las ideas de los neodarwinistas actuales?
Comencemos por las ideas en las que todos coincidimos. Darwin admite que todos los organismos, sean éstos animales domésticos, lombrices de tierra cuyas deposiciones producen «moho vegetal», o percebes (todo lo cual estudió), producen constantemente más descendencia de la que puede sobrevivir. De un modo u otro, nacen, se empollan, se crían y se producen más individuos de los que conseguirán llegar a la madurez. Tan sólo una fracción de los organismos potenciales —en ocasiones tan reducida como uno por cada cien mil, en otras tan cuantiosa como un cincuenta por ciento— llegará a tener descendencia que, a su vez, pueda sobrevivir para tener su propia descendencia. Tan sólo una mínima parte seguirá produciendo descendientes. Darwin reconoció este hecho y le puso por nombre «selección natural». Fundamentalmente, la «selección natural» constituye un modo de expresar el concepto de que la mayor parte de la vida no logra perdurar en el tiempo. De lo que realmente se trata es de «supervivencia diferencial». Podríamos expresarlo del modo siguiente: las poblaciones de todas las especies crecen potencialmente de forma exponencial. En términos de descendencia por generación (o por unidad de tiempo), la tendencia del crecimiento de población se denomina «potencial biótico», refiriéndose al número máximo de descendientes posibles para una especie dada. La selección natural se refiere al hecho de que ese potencial biótico raramente es alcanzado. El potencial biótico de los seres humanos es de veinte descendientes por pareja y generación (o veinte descendientes cada veinticinco años). El potencial biótico del hongo de bosque Alternaría es aproximadamente de 105 (cien mil) esporas por minuto durante seis meses. El del perro salchicha es de once cachorros por camada en tres camadas al año durante diez años o, lo que es lo mismo, 333 cachorros por pareja y generación. Y así sucesivamente. El potencial biótico es específico para cada especie y puede ser expresado en términos del mayor número posible de descendientes nacidos, empollados, criados, germinados a partir de las semillas, o cualquier otra posibilidad de producto de descendencia por cada generación.
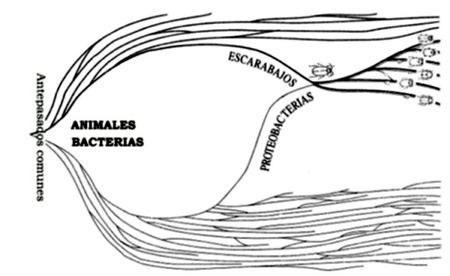
Figura 1.1 Anastomosis subdivisible en la filogenia del escarabajo.
Llegó así a la conclusión de que los organismos están conectados en el tiempo y que todos descienden de antepasados comunes. Denominó a su idea descendencia con modificaciones. Perros y palomas le proporcionaron algunos de sus mejores ejemplos. Observó también que quienes criaban gallinas para comérselas seleccionaban aquéllas con los músculos pectorales más grandes así como que quienes criaban perros para la caza elegían los que tuvieran los sentidos más aguzados. Se dio cuenta de que la selección producía, de este modo, diferencias en la descendencia de forma relativamente rápida, diferencias que, por ejemplo en el caso de los perros, se codificaban como razas distintas.
El concepto básico de Darwin, tal como ha sido modificado, ampliado y verificado en el siglo XX, ha quedado resumido por Ernst Mayr (2001) en su libro What Evolution Is[4]. Esta accesible introducción al gran espectro de la ciencia evolutiva, de la mano de uno de sus participantes en activo, expande el breve resumen que aquí presentamos.
De forma abreviada, el argumento de Darwin es el siguiente: es patente por doquier la abundante producción de descendencia de demasiados organismos, mientras que tan sólo unos pocos sobreviven para reproducirse a su vez. Todos los descendientes varían con relación a sus progenitores. Algunas de estas variaciones son hereditarias. Por consiguiente, en la historia de la vida ocurren variaciones. Sin embargo, lo que tenía perplejo a Darwin era el origen de estas variaciones heredadas. ¿Por qué causa los organismos no eran siempre iguales que sus progenitores o —puestos en ello— que su único progenitor inicial? ¿Cuál era la fuente de la variación evolutiva? Darwin no lo sabía, pero observó que gran parte de las variaciones nunca eran hereditarias: los varones circuncidados, los ratones sin cola y las plantas con hojas amarillentas no daban lugar a niños sin piel en el glande, a ratoncitos sin cola o a plantones amarillentos. Darwin escribió a menudo que tan sólo las variaciones heredadas son importantes para la «descendencia con modificaciones», es decir, para la evolución. La longitud del pelo, en cuanto a variación, carecería de interés puesto que no era heredada. Sin embargo, la longitud máxima que llegara a alcanzar ese pelo, si se le dejara crecer sin límite, podría ser de interés como ejemplo de variación hereditaria.
En resumen, toda población exhibe un grado mensurable de variación, gran parte del cual es absolutamente irrelevante por lo que a la evolución se refiere. Sin embargo, esta misma población exhibe también ciertas variaciones heredadas, relevantes para el cambio evolutivo. Esta clase de variación puede ser detectada y medida en cualquier grupo. Por consiguiente, nuestra pregunta se resume en la misma que Darwin se planteaba a sí mismo: ¿Cuál es la fuente de la variación hereditaria observable?
§. La simbiosis como fuente de variación hereditaria
La idea general entre los neodarwinistas, básicamente zoólogos que en la actualidad se autodenominan “biólogos evolucionistas”, consiste en que la variación heredada deriva de los cambios aleatorios en la química de los genes. Las variaciones heredables son causadas por mutaciones, y éstas son aleatorias. Impredecibles e independientes del comportamiento, de las condiciones sociales, del alimento o de cualquier otro elemento, las mutaciones son cambios genéticos permanentes. A medida que estos cambios genéticos aleatorios van acumulándose con el paso del tiempo, determinan el curso de la evolución. Tal es la visión presentada por la mayor parte de la literatura evolucionista.
Sin duda estamos de acuerdo en que los cambios aleatorios heredables, o mutaciones genéticas, ocurren. Coincidimos también en que estas mutaciones aleatorias quedan expresadas en la química del organismo. La existencia de proteínas alteradas, cuyo origen puede remontarse a mutaciones genéticas en organismos vivos, ha quedado ampliamente demostrada. La diferencia principal entre nuestro punto de vista y la doctrina oficial neo darwinista actual trata de la importancia de la mutación aleatoria en la evolución. Opinamos que la trascendencia de la mutación aleatoria como fuente de variación hereditaria está siendo enormemente exagerada. Las mutaciones —los cambios genéticos en organismos vivos— pueden ser inducidas; es algo que puede hacerse con rayos X o añadiendo compuestos químicos muta génicos en el alimento del organismo. Se conocen numerosas formas de inducir mutaciones, pero ninguna de ellas conduce a la aparición de nuevos organismos. La acumulación de mutaciones no desemboca en el surgimiento de nuevas especies, ni siquiera de nuevos órganos o nuevos tejidos. Si el óvulo o la esperma de un mamífero son sometidos a mutación, ocurrirán efectivamente cambios hereditarios pero, como ya señalara tempranamente Hermann J. Muller (1890-1967), premio Nobel que demostró sobre la mosca de la fruta la capacidad muta génica de los rayos X, el 99,9 por ciento de las mutaciones son dañinas. Incluso los biólogos evolucionistas profesionales tienen serias dificultades para encontrar mutaciones, ya sean inducidas experimentalmente o espontáneas, que contribuyan de forma positiva al cambio evolutivo.
Demostraremos aquí que la fuente principal de variación hereditaria no es la mutación aleatoria, sino que la variación importante transmitida, que conduce a la novedad evolutiva, procede de la adquisición de genomas. Conjuntos enteros de genes, e incluso organismos completos con su propio genoma, son asimilados e incorporados por otros. Es más, demostraremos también que el proceso conocido como simbiogénesis es el camino principal para la adquisición de genomas.
¿Qué es la simbiogénesis y cómo está relacionada con la simbiosis? La simbiosis es, simplemente, la vida en común de organismos distintos entre sí. Definida originalmente por Heinrich Anton de Bary (1831-1888), la simbiosis consistía en la cohabitación de «organismos de denominación distinta». Las simbiosis son asociaciones de largo recorrido. Organismos de tipología distinta se unen y dan pie a un tercer organismo. Esta fusión no es aleatoria; las relaciones simbióticas ocurren bajo condiciones del medio específicas. En algunas de estas asociaciones, una de las partes alimenta a la otra incluso a costa de su propio detrimento o de su muerte. Esta clase de asociaciones explotadoras se conocen como “parasitarias” o “patogénicas”. Tienden a ser extremadamente sensibles al estrés medioambiental. El parásito que invariablemente y de forma virulenta mata a su anfitrión se mata a sí mismo. Con el tiempo y las circunstancias, la naturaleza de las asociaciones tiende a cambiar. Las relaciones que aquí nos interesan principalmente son la coexistencia modulada entre anteriores predadores, así como entre patógenos y sus anfitriones, sus cobijos y sus fuentes de alimento. En la medida en que, con el paso del tiempo, los miembros de dos especies distintas responden a la presencia del otro, las relaciones explotadoras pueden llegar a transformarse en convivencia, hasta el punto en que ninguno de los dos organismos pueda ya existir sin el otro.
La simbiosis estable a largo plazo que desemboca en cambio evolutivo recibe el nombre de “simbiogénesis”. Estas asociaciones, verdaderas fusiones biológicas que se inician en forma de simbiosis, constituyen el motor de la evolución de las especies. Veamos cómo esclarece esta cuestión un ejemplo muy específico de simbiogénesis en organismos vivos. Vamos a presentarte a los animales fotosintéticos, verdaderos híbridos entre vegetal y animal. Las babosas, esos conocidos moluscos sin cáscara que se comen las plantas de tu jardín, tienen unos parientes fotosintéticos completamente verdes. Los antepasados de esas babosas verdes comieron años a ciertas algas verdes que no llegaron a digerir, por lo que éstas pasaron a formar parte de los tejidos del animal, quedándose allí. Todos los miembros de esta especie (por ejemplo la Elysia viridis) son siempre verdes. Estas babosas subacuáticas no tienen que preocuparse por la comida. Desde que llegan a la etapa adulta, dejan de comer. Para alimentarse se arrastran hasta la orilla donde, igual que a las plantas, a los miembros de esa nueva familia de animales verdes les basta con tomar el sol. Son permanente y discontinuamente distintos de sus antepasados grises, comedores de algas. Tal vez esta adquisición, utilización y herencia permanente de genomas enteros de alga pueda parecer anecdótica, pero, de hecho, ha sucedido en numerosas ocasiones a lo largo de la historia de la evolución. La existencia de distintos linajes de animales verdes ha quedado acreditadla, al menos cuatro o cinco veces, a través de videos o de publicaciones específicas. Los animales verdes nos proporcionan ejemplos gráficos de simbiosis que desembocan en simbiogénesis.
El liquen, ese verdor en forma de planta que crece sobre las rocas y los troncos de los árboles, es otro buen ejemplo. La mayoría de biólogos no se percatan de que son schwendeneristas. Un schwendenerista es un científico —habitualmente un botánico— que cree que los líquenes no son plantas. La cuestión schwendeneriana tuvo su apogeo a finales del siglo XIX. Simón Schwendener (1829-1919) explicó, en un tratado largo y complejo escrito en alemán, la naturaleza compuesta de las manchas de líquenes y por qué ningún liquen era una planta. Sin embargo, todos los investigadores modernos aceptan el origen simbiogenético de los líquenes. Se sabe ya que una cuarta parte de la totalidad de los hongos documentados están “liquenizados”, es decir, necesitan vivir fotosintéticamente en asociación con algas o con cianobacterias. Este hecho abarca por sí solo unas 25.000 especies de líquenes. Los organismos fusionados actúan como una única entidad: producen tejidos altamente organizados y estructurados de tallos, hojas y protuberancias bulbosas. Las células de hongos y de algas (o de cianobacterias) de los líquenes se propagan juntas. Ningún biólogo riguroso se atrevería hoy a afirmar que algún liquen es una planta verdadera. Todos están de acuerdo en que los líquenes parecen plantas y foto sintetizan como tales, aun cuando un análisis morfológico superficial demuestre que difieren fundamentalmente de ellas.
Los líquenes nos proporcionan un ejemplo característico de simbiogénesis. Es más, el individuo liquen es algo diferente de sus dos componentes. No es ni un alga verde o una cianobacteria, ni un hongo. Es un liquen. Los líquenes, novedades evolutivas surgidas por medio de la adquisición de genomas de alga o de cianobacteria, tomaron su propio camino y exhiben características distintas a las de sus antepasados. Aunque estudiados tradicionalmente dentro de la botánica, los líquenes han sido fundamentales para los conceptos de simbiosis y simbiogénesis en el pensamiento evolutivo, a pesar de lo cual su naturaleza simbiótica ha hecho que se los considerara como fenómenos evolutivos marginales. Tal vez hayan sido aceptados como un ejemplo del poder de la simbiogénesis para generar novedad evolutiva, debido únicamente a que ambos asociados son del mismo tamaño. Tanto las algas como los hongos pueden observarse con facilidad, simplemente con la ayuda de un microscopio de pocos aumentos, de modo que no es posible estudiar las unas sin estudiar simultáneamente los otros. En cambio, en algunos animales verdes (como en el caso de la especie de lombriz plana Convoluta roscoffensis) los respectivos tamaños de los componentes difieren enormemente. La lombriz mide centímetros, mientras que los diminutos organismos fotosintéticos —las algas— son microscópicos. Tales discrepancias de tamaño hacen que, tanto la simbiosis como la correspondiente simbiogénesis, resulten menos evidentes.
La actitud habitual consiste en decimos: «Bueno, la simbiosis es aceptable para la evolución de esas cosas microbianas que os interesan a vosotros (como si tuviésemos alguna clase de extraña fijación con los microorganismos), pero no creemos que la simbiogénesis sea un mecanismo evolutivo verdaderamente importante para los organismos “superiores” (principalmente mamíferos), que son los que nos interesan a nosotros realmente». Sin embargo, este libro rebosa de ejemplos en los que la simbiogénesis sirve como fuente de novedad evolutiva en animales bien conocidos, como por ejemplo las vacas. Según sir David C. Smith (Smith y Douglas, 1987), las vacas son «tanques de fermentación de ciento cincuenta litros sobre cuatro patas». Las vacas ingieren hierba, pero nunca la llegan a digerir porque no pueden descomponer la celulosa. La digestión ocurre en el interior de la vaca gracias a sus simbiontes microbianos alojados en el rumen o panza, una cavidad especial del estómago —en realidad un esófago superdesarrollado— que ha ido evolucionando con el tiempo. No existen vacas sin rumen; las vacas o los toros privados de sus simbiontes microbianos no pueden vivir.
Los cambios aleatorios en la base de ADN juegan, sin duda, un papel en el proceso evolutivo. Son como errores de imprenta que se multiplican en cada ejemplar del libro. Raramente contribuyen a clarificar o ampliar el sentido del texto. Tales pequeños cambios aleatorios son casi siempre inconsecuentes —o incluso dañinos— para el conjunto de la obra. No es que estemos negando aquí la importancia de las mutaciones. Únicamente insistimos en que, siendo tan sólo una pequeña parte de la saga evolutiva, la mutación ha estado siendo dogmáticamente sobrevalorada. La parte mucho mayor de la historia de la innovación evolutiva, correspondiente a la unión simbiótica de organismos de linajes distintos, parecida —por seguir con la analogía— a la fusión de textos mediante el plagio o la antología, ha estado siendo sistemáticamente ignorada por los autoproclamados biólogos evolutivos (Sapp, 2002).
En el árbol filogenético corriente, la adquisición de genomas heredables puede ser descrita como anastomosis o fusión de ramas. La principal proposición que aquí formulamos, a saber, la fusión de linajes evolutivos, es en ocasiones descrita como una alternativa al darwinismo clásico. Sin embargo, la adquisición simbiogenética de nuevos rasgos por medio de la herencia de genomas adquiridos constituye más bien una extensión, un refinamiento y una amplificación de la idea de Darwin. Esta evolución requiere nuevos procesos de pensamiento, hacen falta nuevas metáforas que reflexionen sobre las asociaciones permanentes. La simbiosis, la unión, la fusión de cuerpos, no pueden ser reducidas a la sustitución de la “competición” por la “cooperación” como principal fuerza motriz de la evolución. En última instancia, un término antropocéntrico como “competición” está obviamente fuera de lugar en el diálogo científico. En lugar de ello nos conviene proponer una búsqueda en las ciencias sociales para dar con términos nuevos que reemplacen a las viejas y caducas metáforas sociales de Darwin. Si la supervivencia se debe efectivamente a la simbiosis, más que a las sobrevaloradas pugnas competitivas entre especies, ¿cuáles son entonces las consecuencias de ello para los no biólogos interesados en la evolución?
En la presente obra hemos evitado escrupulosamente utilizar los términos “competición” y “cooperación” para describir fenómenos biológicos o evolutivos de otra índole. Estas palabras tal vez resulten adecuadas para el baloncesto, la industria informática o las instituciones financieras, pero en cuanto a la evolución, pintan con una brocha demasiado gorda. Con demasiada frecuencia olvidan las complejas interacciones que se dan entre los seres vivos, entre los organismos que cohabitan. La competición implica un acuerdo previo, una serie de acciones que se ciñen a determinadas reglas. En el juego de la vida real, sin embargo, las “reglas” —basadas en la química y en las condiciones del medio— cambian con los jugadores. Para competir, por ejemplo en equipos opuestos, las personas tienen que cooperar básicamente en cierta medida. “Competición” es un término con significado científico limitado, habitualmente carente de referencia alguna a las unidades en las que pueda ser medida. ¿Cómo evalúan la lombriz verde o el hongo liquen su estatus competitivo? ¿Añadiendo puntos a su calificación, o en dólares o en francos suizos? Ciertamente que no. ¿Cuáles son entonces las unidades de competición? Si nos preguntas por las unidades de medición de la biomasa te podemos responder en gramos u onzas. Si nos interrogas acerca de la forma de medir el potencial lumínico o biótico te responderemos en luxes o bujías y en número de descendientes por generación. Pero si nos preguntas «cuáles son las unidades de competición» te contestaremos que tu visión no es científica. Términos en boga como “competición”, “cooperación”, “mutualismo”, “beneficio mutuo”, “costes energéticos” y “ventaja competitiva”, han sido importados de las empresas humanas e impuestos a la ciencia desde la política, el mundo de los negocios y la reflexión social. Toda la panoplia de términos neodarwinistas refleja un mismo error filosófico, un ejemplo del siglo XX del fenómeno que acertadamente Alfred North Whitehead denominara «la falacia de la concreción mal atribuida». La terminología de la mayoría de evolucionistas modernos no es tan sólo falaz, sino también peligrosa, puesto que conduce a la gente a creer que entiende de la evolución de la vida cuando, en realidad, está confundida y mal informada. El «gen egoísta» es un buen ejemplo de ello. ¿Qué es ese gen egoísta de Richard Dawkins? Para comenzar, un gen nunca puede ser un ego. Un gen aislado es tan sólo un fragmento de ADN capaz de tener una función. El gen por sí sólo no es nada; aunque lo conservemos en un congelador o en una disolución salina, el gen aislado nunca tendrá actividad por sí mismo. No hay vida en un gen, no es un ser, en ningún caso puede reunir los criterios mínimos para ser una entidad, un sistema vivo. Ha llegado el momento de que la biología seria deje de lado términos como competición, cooperación y genes egoístas, para reemplazarlos con otros tales como modalidades metabólicas (quimioautotrofia, fotosíntesis), relaciones ecológicas (epibiotas, polinizadores) y cantidades mensurables (luz, calor, fuerza mecánica). Demasiadas de las metáforas evolutivas en uso no son más que dicotomizaciones superficiales, procedentes de engañosas claridades de lenguaje. Lejos de producir comprensión científica, la impiden.
¿No sería pues mejor para la sociedad que adoptáramos metáforas simbióticas en lugar de competitivas? Pues no, lo mejor para la sociedad no es que reemplacemos una ristra de metáforas sobre simplificadas por otra, sino que avancemos en la exactitud de la comprensión científica. Ciertamente, los organismos compiten entre sí de diversas formas por espacio y alimento. No obstante, esta pugna (o competición) entre miembros de la misma especie no conduce a la emergencia de nuevas especies. Para ello es necesaria una fuente de novedad genética, que habitualmente es la simbiogénesis. Tan sólo sobrevive una pequeña porción de cualquier población. ¿No es eso entonces competición? ¿No es eso una «lucha por la supervivencia»? No realmente. Cuando Darwin hablaba de lucha, se refería a la tendencia de todo organismo a crecer, a reproducirse y a tratar de dejar tras de sí su propia descendencia. No se refería a que Dios luche con ángeles o que las concubinas del rey se peguen puñetazos unas a otras. En el sentido de la tendencia de algunos organismos —pero no de todos— a dejar descendencia, el término darwiniano adecuado es selección natural. Su “lucha” consiste en el crudo hecho de que, como regla de vida, el potencial biótico nunca es alcanzado. Tan sólo unos pocos de entre nosotros producen descendencia que seguirá produciendo descendencia que, a su vez, produzca también descendencia fértil. Llamar “competición” a la tendencia general a dejar descendencia tras de sí, como suelen hacer los biólogos, es un error.
Como especie elevadamente social que somos, preocupada frecuentemente por las relaciones entre nosotros mismos, tendemos a olvidar nuestras relaciones con las demás especies. Biológicamente, por ejemplo, no existe cosa tal como una simbiosis entre una madre y su hijo aún no nacido. La simbiosis es siempre una estrecha relación física entre organismos de distinta especie. Los humanos tenemos una relación simbiótica con los ácaros de las pestañas. La mayoría de nosotros ignoramos el hecho de que vivimos simbióticamente con estos organismos, o con las bacterias de las axilas o de los intestinos, o con las espiroquetas del tejido de nuestras encías. Pero lo cierto es que tenemos esos ácaros en las pestañas y esas bacterias en las axilas y en los intestinos. No prestamos atención a nuestras inmensas poblaciones de espiroquetas orales, hasta el día en que nos sangran las encías o nos duelen las muelas. Todos nosotros albergamos en nuestros intestinos bacterias que producen vitamina K y B que son absorbidas por las paredes intestinales. Parece que nos cuesta aceptar a nuestras bacterias simbióticas, aun cuando podamos verlas perfectamente a través del microscopio electrónico. Disfrutamos todos de relaciones silenciosas e inconscientes con microbios.
Nadie de nosotros tiene, en cambio, una relación simbiótica con su suegra, con su padre o con su hijo adolescente. ¿Por qué? Porque todas las personas pertenecen a una misma especie, la del Homo sapiens. “Simbiosis” es un término de ecología[5] que describe a dos o más organismos de especies distintas, en contacto físico prolongado. Incluso las abejas y las flores, a pesar de su íntima asociación en la ecología de la polinización, no tienen entre sí una relación simbiótica. Los ácaros de las pestañas y los seres humanos son, en cambio, simbiontes. El olor que emana de tus calcetines procede de algunas de las bacterias y de los hongos que viven entre los dedos de tus pies, donde tienen su hábitat específico. Se estima que el diez por ciento del peso en seco de cualquier persona corresponde a simbiontes. La mayor parte de estos simbiontes, por cuanto a peso se refiere, se compone de los numerosos tipos de bacterias que medran en el colon o intestino grueso. Nuestra relación con esos microbios es de asociación:

En resumen, gran parte de la terminología de los evolucionistas (no de los extraños darwinistas) modernos debería ser descartada. Tanto en la cultura popular como particularmente en la subcultura científica de la «biología evolutiva», la terminología no es científica, sino engañosa hasta el punto de ser destructiva. Los críticos sinceros del pensamiento evolucionista que se atreven a señalar los problemas del dogma de los biólogos, junto con su terminología indefinible, son a menudo desdeñados como si de fundamentalistas religiosos o xenófobos se tratara. Sin embargo, la parte de la tesis defendida en este libro, que insiste en que dicha terminología (ver tabla 1.2) interfiere con la verdadera ciencia, demanda un debate abierto y profundo acerca de la veracidad de las afirmaciones formuladas por los evolucionistas zoocéntricos.
§. El mito del individuo independiente
Tal vez te sorprenda la sugerencia de que los organismos no son tan individuales como solemos creer. La idea de que en realidad somos montajes ambulantes, seres que han integrado diversas clases de organismos extraños, de que cada uno de nosotros es una especie de comité anárquico, abre la puerta a demasiadas especulaciones retadoras. Por ejemplo, cuando el comité cae enfermo, ¿es un único animal el que ha enfermado, o esa enfermedad es más bien un desajuste entre los miembros que lo componen? Imaginamos que los microbios patógenos nos atacan pero, si resulta que estos patógenos forman parte de ese comité que nos hace ser lo que somos, ¿no será la salud una cuestión de relaciones ecológicas entre los miembros del comité, más que de resistencia a las invasiones procedentes del exterior? Sin duda que sí.
Como cualquier otro organismo, los seres humanos vivimos inmersos en comunidades ecológicas. Cuando, como individuos, sentimos que nos estamos derrumbando, ello se debe probablemente a que, efectivamente, ese edificio compuesto de múltiples elementos que somos se está viniendo abajo. Cada persona, cada perro, cada árbol, se compone de numerosas partes vivas diferentes que pueden ser detectadas e identificadas. Las relaciones entre las partes vivas que nos componen son absolutamente cruciales para nuestra salud y, por consiguiente, para nuestra felicidad. El “individuo” absolutamente auto contenido constituye un mito que necesita ser reemplazado por una descripción más flexible. Sin embargo, los simbiontes del ser humano son difíciles de estudiar, y ello por numerosas razones: complejidad debida a la disparidad de tamaños, incapacidad de llevar a cabo experimentaciones con la herencia humana, política y prejuicios sociales. Por todo ello, los líquenes nos proporcionan más facilidades para el estudio de los simbiontes. Si privamos de luz a ciertos líquenes, el miembro fotosintético o fotobionte (habitualmente una cianobacteria como la Nostoc o un alga verde como la Trebouxia) no puede vivir en la oscuridad. Entonces el hongo suele crecer sin parar, digiriendo a su anterior socio. Si, por el contrario, sumergimos al liquen durante largo tiempo bajo el agua pero con luz, el hongo termina por ahogarse, mientras que el alga crece continuamente. Por consiguiente, los líquenes son organismos compuestos que requieren luz variada: no pueden sobrevivir en condiciones persistentes de todo-luz o nada-luz. Del mismo modo, tampoco pueden vivir en condiciones de sequedad o de humedad absolutas, sino que precisan de un ciclo entre ambos extremos. Son estos ciclos, estas alternancias entre humedad y sequedad y entre luz y oscuridad, las que mantienen la composición viva, el liquen con apariencia de individuo. Para que la mayoría de seres vivos puedan persistir y propagarse son necesarias determinadas ratios de cambios.
Es necesario que comencemos a pensar en los organismos en términos de comunidades, de colectivos. Las comunidades son entidades ecológicas.
Para ir más allá de los animales, hay que pensar en las plantas. Plantamos sus semillas o sus esquejes en el suelo del huerto y nos maravillamos al verlas hacer su parte. La mayoría de las raíces viven en la rizosfera, esa zona ecológica en la que medran numerosos organismos distintos que crecen y metabolizan al mismo tiempo. Algunos de sus moradores proporcionan nutrientes a eso que vemos como una planta que es, generalmente, lo que está por encima del suelo. Pero, al igual que los animales, las plantas son también confederaciones de organismos de distintas clases, otrora separados. Como veremos, se sabe que las simbiosis en las raíces, en las hojas, e incluso en los tallos, han dado lugar a nuevas formas de vida vegetal, siendo tal vez responsables del origen de esos crecimientos, en su día monstruosos, sin los que la humanidad nunca hubiera evolucionado en lo que hoy conocemos como tal: los frutos.
El libro ¿Qué es la vida? (Margulis y Sagan, 2000) incluye una fotografía tomada a distancia por Connie Barlow de un bosque de álamos en Colorado. Cualquiera puede contar los árboles. Hay centenares, si no miles de ellos. Sin embargo, y aunque consta de numerosas partes, ese bosque es en realidad un único organismo. Bajo la superficie del suelo, el “árbol” es uno continuo. Constituye una estructura interconectada con múltiples tallos que surgen del suelo, cuestionando nuestra noción común de árbol como organismo aislado. Ese “individuo”, cuyas raíces son enteramente continuas, se extiende lateralmente durante kilómetros, se hunde varios metros en el suelo y se eleva en el aire en vertical. Se cree que esta plantación de álamos es uno de los mayores “organismos” vivos de nuestros días.
Nuestro concepto de proceso evolutivo necesita incorporar íntegramente a los vegetales. Como ya hemos señalado, el problema con los “biólogos evolucionistas” modernos consiste en que sus ejemplos derivan casi siempre de los humanos o de otros animales, particularmente mamíferos foráneos. En escasas ocasiones, el catálogo se extiende a la mosca de la fruta o a algún otro insecto que les resulte de utilidad para su explicación de la evolución. Cuando hablan de «organismos inferiores» suelen referirse a los animales que no son mamíferos. Como zoólogos que son, tienden a desconocer el mundo microbiano, ignorando a menudo a las bacterias, a los hongos y a la miríada de esos otros microbios “mayores” denominados protistas. Los zoólogos tienden a estudiar muy poca botánica, escasa protistología y ninguna bacteriología. Escriben sobre “individuos” pero habitualmente se refieren a personas, mascotas y animales de granja o de zoo. Ocasionalmente tal vez incluyan algún vegetal agrícola en sus análisis, pero a menudo lo hacen de forma limitada y científicamente inadecuada. Aunque el zoocentrismo pueda resultar apto para la clase de mamíferos empleados para la cría de poblaciones mayores, no deja de ser una burda aproximación —una especie de alucinación apolínea— de lo que realmente sucede en la vida como un todo.
Sigue con nosotros la pregunta decimonónica de si existe o no un progreso evolutivo. ¿Debe entenderse como tal la complejidad creciente a gran escala evolutiva?
La evidencia de expansión evolutiva es fácil de demostrar. Abundan en los registros fósiles las pruebas de la expansión de nuevas formas de vida. La vida —incluyendo por supuesto a la vida humana— gusta de estar donde el agua, en forma de océano o lago, está en contacto tanto con el aire como con el suelo. Todos sabemos, por el valor inmobiliario de esos suelos, que las franjas costeras son lugares apetecibles. A la vida le gustan los hábitats donde coinciden agua, tierra y aire. La mayor parte de la vida ocurre a pocos milímetros de las superficies del agua y del suelo.
Aparentemente, la vida evolucionó a partir de su propagación en la costa marina, expandiéndose tan sólo después a las regiones polares, a los lagos de alta montaña y a los abismos oceánicos. Únicamente a partir de la era cenozoica, que comenzara hace 64 millones de años, comienzan los animales y las algas a dejar registros fósiles en el alto Ártico y en la Antártida (para una escala geológica temporal simplificada ver tabla 9.1). El continuo núcleo central de la vida sobre el planeta se ha expandido y ha extendido su espectro. ¿Es eso progreso? Se han hallado fósiles de reptiles que exhiben más vértebras y mayor complejidad morfológica que cualquier otro ser vivo conocido. Habida cuenta de su extinción, su testimonio contradice el concepto mismo de progreso evolutivo. El propio término “progreso”, con sus connotaciones morales, denota una cantidad compleja inmensurable e inestimable. Los descendientes de esos reptiles perdieron complejidad, son parientes simplificados de sus antepasados, pero tampoco podemos decir por ello que hayan “retrocedido”. Simplemente evolucionaron hasta tener menos vértebras, eso es todo. Que algún progreso direccional nos haya encumbrado a nosotros, Homo sapiens, a una supuesta cima olímpica, constituye una idea absolutamente insostenible. A medida que la Tierra se iba cubriendo de más y más vida, ésta se expandía, pero que “progresara” o no es un tema cuestionable. Como veremos en el capítulo siguiente, el aparente progreso de la vida se entiende mejor en el contexto de su conformidad con la segunda ley de la termodinámica. En dicho capítulo seguiremos el rastro de la vida desde el Precámbrico hasta el presente, manteniendo nuestra atención sobre la pregunta, « ¿cómo evolucionan las nuevas especies?» Remoldearemos los conceptos de “progreso evolutivo” en términos de la nueva termodinámica, que unifica e integra, de forma distinta pero complementaria a la genética y a la biología molecular, la ciencias de la física y de la biología. Contenido:
Darwinistas de última horaLos 1.200 ejemplares publicados inicialmente de El origen de las especies por medio de la selección natural, o la preservación de las razas aventajadas en la lucha por la vida de Charles Darwin, se vendieron al librero el mismo día de su publicación, el 24 de noviembre de 1859. Los lectores eran conscientes de los rumores que circulaban: el autor del popular Viaje del Beagle iba a explicar de qué modo el ser humano descendía del mono, por qué la risa y las muecas eran expresiones faciales comunes a reyes y esclavos, y de qué modo se había retirado Dios Todopoderoso de su trabajo como productor, presidente y director general ejecutivo del mundo conocido. Por fin, el aislado académico, que había ido construyendo su teoría laboriosamente a lo largo de tantos años, tras su empleo juvenil como naturalista en el Beagle, iba a contar toda la verdad. Todos sabían que aquel Darwin era cuidadoso y merecedor de toda confianza y que, habida cuenta de que la fortuna de su familia y de la de su esposa, Emma Wedgwood, le permitían dedicarse por completo a la búsqueda de la verdad, el contenido de su nuevo libro no podía ser más que exacto y fiable.
Herencia y salud
Células, microbios, propágulos y entidades autopoiésicas
Termodinámica, «propósito» y «progreso» evolutivo
Lo más probable es que el libro fuera mucho más comprado que leído. Tras un siglo de exégesis por la sabiduría establecida y numerosas traducciones al lenguaje común, las autoridades competentes han decretado que las intuiciones de Darwin fueron tan numerosas como valiosas. Sin embargo, el contenido de la aludida obra está repleto de dudas, de contradicciones y de posibles prevaricaciones. En ella, el autor deja claro que el ser humano, como todos los demás seres vivos, ha sobrevivido hasta el presente precedido por un historial inmenso y sobrecogedor. Ningún dios es responsable de la construcción del ser humano, como tampoco de la del tomate. Ninguna otra forma de vida fue tampoco creada por separado en siete días o menos. No obstante, Darwin, probablemente por razones principalmente de índole política, dejaba la puerta abierta a la posibilidad de un origen divino de la vida en su inicio.
Sorprendentemente, tras todas las explicaciones acerca de la «grandiosidad de esta nueva visión de la vida» (una de las frases de Darwin en su gran obra), queda claro que, en las 500 páginas de tipografía menuda apretada, la obra en cuestión se olvida por completo de su propio título —el origen de las especies—, que queda abandonado y relegado al olvido. Como señalara tan acertadamente el biólogo australiano George Miklos.
La «lucha por la supervivencia» ha sido aceptada incuestionadamente por generaciones de biólogos evolucionistas, que citan el Origen de las especies como si de las Sagradas Escrituras se tratara, a pesar de que el origen de las especies sea, precisamente, de lo que la obra de Darwin no trata.Tanto para quienes hayan hojeado el libro, como para quienes hayan leído los otros numerosísimos trabajos de su autor, o sepan de él por persona interpuesta, lo que Charles Darwin estableció en última instancia en su famoso libro, para mayor satisfacción de sus coetáneos científicos y de sus seguidores, fue una idea fundamental que sigue siendo enteramente válida en nuestros días. Toda especie viva desciende de otra anterior relacionada con ella. Toda forma de vida, haya sido o no creada por una supuesta deidad en su inicio, está conectada a través del tiempo con otras formas de vida preexistentes, probablemente similares a ella. Hoy en día, con la mejor comprensión de la evolución cósmica y de la química de los orígenes de la vida de que disponemos, la necesidad de un origen divino debe remontarse aún más, hacia el misterioso nacimiento del cosmos en el Big Bang.
Darwin demostró claramente que los seres vivos “engendran” descendientes que, inevitablemente, difieren ligeramente de sus progenitores. Observó que muchas de estas diferencias identificables son heredadas. Puede comprobarse la transmisión de rasgos, a menudo distinguibles aunque ligeramente cambiados, en cachorrillos de perro, en potrillos, en pollitos, en temeros, en hijos y en nietos. Darwin no hizo más que divulgar un hecho que todos podían ver por sí mismos: del gran número de descendencia que llega a surgir de semillas, huevos y úteros, tan sólo un pequeño número sobrevive y consigue engendrar su propia descendencia. Tal vez sean muchos —bellotas y cachorros por igual— los que estén al principio alrededor de la madre pero, si se sigue el rastro de su vida como individuos, se comprueba que son bien pocos los que consiguen sobrevivir, madurar y dejar descendencia fértil. Por lógica —argumentó Darwin— los sobrevivientes tienen que tener rasgos que sean más favorables a la supervivencia en ese entorno concreto, que sus congéneres que no lograron sobrevivir. Como ya ha quedado dicho, es a ese proceso de supervivencia y reproducción diferenciales al que Darwin aludía con la denominación de «selección natural».
Esa «selección natural» de Darwin no implicaba de ningún modo ni un lúgubre, anciano y poderoso morador de un cielo tormentoso, ningún «Seleccionador Natural». Su expresión denotaba la supervivencia y la reproducción de unos pocos elegidos, parientes de un prodigioso número inicial de descendientes. Darwin admitía que, por sí sólo, ese proceso de selección natural no parecía ser capaz de crear novedad, limitándose simplemente a eliminar, del inmenso catálogo de diferentes organismos presentes en la Naturaleza, aquellos individuos incapaces de reproducirse. Pero, ¿dónde se originaba esta variación intrínseca y heredada, postulada por Darwin?
Parecería como si Darwin quisiera hacernos creer que el concepto entero de evolución comienza con él. Consistentemente deja de hacer honor a su vital abuelo paterno, Erasmus Darwin, médico y poeta progresista cuya contribución en Zoonomia (1794-1796) acerca de la selección natural, no parece haber sido tomada demasiado en serio por su nieto. Jean Baptitste Lamarck (1774-1829) fue el primer naturalista moderno en publicar un gran Corpus de bibliografía argumentando la evolución de toda forma moderna de vida a partir de antecesores ancestrales. En los círculos anglófonos, se habla de Lamarck como del francés responsable de una contribución negativa a la ciencia, con su afirmación errónea de que las características adquiridas por un animal o una planta pueden ser heredadas por los descendientes del adquiriente.
La «herencia de características adquiridas», expresión inseparable ya del nombre de Lamarck, se conoce como “lamarckismo” y se equipara con error. Sin embargo, y al igual que Lamarck, el propio Darwin se debatió con el problema de la fuente primigenia de la variación heredable, llegando también a conclusiones erróneas. Al parecer se prefiere olvidar que, como señala Mayr en su libro (1982), Darwin acabó por inventarse una explicación lamarckiana —su hipótesis “pangenética”— para explicar el origen de las variaciones heredables. Según esta teoría las “gémulas”, supuestas partículas de las que todo ser vivo estaría dotado y sujetas a la experiencia durante la vida de sus portadores, mandan representantes a la siguiente generación. El punto de vista de Darwin, difícilmente diferenciable del de Lamarck, constituye una declaración formal en favor de la «herencia de características adquiridas». En última instancia, sin embargo, Darwin se equivocó en el origen de esas “cosas”, de esos “mutantes”, de esas “variaciones heredables”. Simplemente, lo desconocía.
§. Darwinistas de última hora
Partiendo de Inglaterra después de su muerte y habiéndose extendido por Alemania y los Estados Unidos en gran parte del mundo alfabetizado, el legado de Darwin sigue hoy en boga en infinidad de textos y aulas como —en nuestra opinión— el sistema de creencias idiosincrásico de la mayor parte de los “evolucionistas modernos”. De forma abreviada este sistema diría lo siguiente: por medio de la selección natural, todos los organismos derivan de antepasados comunes. Las mutaciones aleatorias (los cambios heredables) aparecen en los genes —en el ADN— de los organismos. En comparación con los demás, los mejores “mutantes” —individuos que soportan las mutaciones— son naturalmente seleccionados para sobrevivir y perdurar. Los descendientes poco adecuados —frecuentemente denominados “no aptos”— mueren. La “aptitud” —término técnico— se refiere al número relativo de descendientes que cada individuo deja en la generación siguiente. Por definición, la mejor aptitud produce el número de descendientes más elevado. Las variaciones mutantes dejan así más descendencia y la población evoluciona, es decir, cambia con el paso del tiempo. Cuando el número de cambios en la descendencia se acumula hasta proporciones reconocibles en poblaciones geográficamente aisladas, emergen gradualmente nuevas especies. Cuando se acumula un número suficiente de cambios en la descendencia, aparecen gradualmente taxones superiores, en el sentido de más inclusivos. A lo largo de períodos geológicos de tiempo, las nuevas especies y los taxones superiores (géneros, familias, órdenes, clases, filums, etc.) son fácilmente distinguibles de sus antepasados.
Estamos de acuerdo en que muy poca de la descendencia potencial consigue sobrevivir y tener descendencia, así como en que las poblaciones cambian con el paso del tiempo y que, por consiguiente, la selección natural es de una importancia crítica en el proceso evolutivo. Sin embargo, la explicación darwiniana sobre la totalidad de la evolución es una verdad a medias, cuya carencia de poder explicativo sólo puede ser compensada con la ferocidad religiosa de su retórica. Aun cuando las mutaciones aleatorias hayan alterado el decurso de la evolución, su influencia se debe principalmente a la pérdida, a la alteración y al refinamiento. Una mutación puede conferir resistencia a la malaria, pero también puede convertir a los glóbulos rojos sanos en los deficientes transportadores de oxígeno de la anemia falciforme. Otra mutación convierte a un espléndido recién nacido en víctima de la fibrosis quística, o de una diabetes prematura. Es una mutación la que le impide volar a la mosca de la fruta de ojos rojos. Sin embargo, ninguna mutación ha conseguido que aparezcan alas, que nazca un fruto, que brote un tallo o que surja una uña. En general, las mutaciones tienden pues a generar enfermedades, deficiencias o muerte. No existe, en toda la vasta literatura sobre el cambio hereditario, prueba alguna que demuestre de forma indiscutible que, ni siquiera en el caso de poblaciones geográficamente aisladas, la mutación aleatoria conduzca a la aparición de nuevas especies. Siendo ello así, ¿cómo aparecen las especies? ¿Cómo llegaron las coliflores a descender de unas pequeñas plantas mediterráneas en forma de col? ¿Cómo evolucionó el cerdo a partir del jabalí salvaje?
Sucesores de Darwin han regresado a las islas volcánicas del archipiélago de las Galápagos, frente a las costas del Ecuador, para observar a la evolución en acción. Tal vez el caso real más convincente de estudio del cambio evolutivo sin interrupciones sea el trabajo realizado por los profesores Rosemary y Peter Grant, exactamente con los mismos pinzones que Darwin y sus sucesores hicieran famosos. La escasez de otros pájaros en aquellos remotos lugares, junto con la severidad de sus condiciones ambientales, condujo de algún modo a una rápida y rampante especiación partiendo de antepasados comunes. Como describe Jonathan Wiener en su libro The Beak of the Finch (1999), el estudio de la variación y de los cambios en estos pájaros, aislados del continente sudamericano durante un millón de años, nos proporciona el mejor punto de vista tradicional posible acerca del modo en que debería funcionar el proceso de especiación.
Los ornitólogos actuales reconocen trece especies de pinzones de las Galápagos, que clasifican en cuatro géneros. Los seis pinzones terrestres, que tienden a volar únicamente como parte de sus ritos de apareamiento, pasan el resto de su tiempo saltando por ahí. Estas aves terrestres se agrupan en los géneros Geospiza: G. fortis (fuerte) o pinzón terrestre mediano, G. magnirostris (pico grande) o pinzón terrestre grande, G. difficilis o pinzón terrestre de pico agudo (el más difícil de distinguir), G fuliginosa (el que huye) también denominado pequeño pinzón terrestre, y G conirostris o gran pinzón del cactus, denominado así probablemente a causa de su masivo pico en forma de cono, apto para quebrar grandes semillas de cactus, cuyo nombre común es peligrosamente parecido al pinzón común del cactus o G. scandens.
Las otras siete especies de pinzones se dividen en tres grupos: los que viven en los árboles alimentándose de frutos e insectos, los arbóreos estrictamente vegetarianos y los que habitan entre las ramas de los árboles y muestran “evolución convergente”, puesto que cantan, se comportan y se alimentan de forma tan parecida a las currucas, que al principio se les tomó por tales.
Desde el año 1978, los Grant han estado acampando en la isla Genovesa, cerca de una laguna sin tiburones ni sanguijuelas que lleva por nombre el del afamado Darwin. Allí han estado tomando datos sobre cantos, patas, huevos y picos de pinzones. La totalidad del archipiélago de las Galápagos sufrió una extremada sequía en 1977, año en que probablemente no cayera sobre aquellas islas ni una sola gota de lluvia. El árido verano de 1985 fue igualmente seco. Sin embargo, en el año de “el Niño” de 1982-1983, probablemente la temporada más húmeda de la historia del archipiélago, el registro de precipitaciones rozó los 200 milímetros, con efectos drásticos sobre los pájaros y sus fuentes de alimentación. El fluir de lodos volcánicos determinó una explosión de vegetación. Las parras se enroscaban en los postes de las tiendas de campaña y el crotón floreció siete veces en lugar de una. El número de semillas sobre el suelo fue siete mayor que el del año anterior. Las orugas multiplicaron su población por cinco y estaban más gordas que de costumbre. Los grandes cactus, sin embargo, no lograron sobrevivir. Los héroes del desierto quedaron inundados y desbordados por las pequeñas plantas. En unas orgías nunca antes vistas, los pájaros copulaban, ponían sus huevos y volvían a copular. Este amplio cambio ambiental de seco a húmedo y de nuevo a seco generó un festín de números para los científicos. Las poblaciones experimentaron grandes fluctuaciones, desde los apuros de los grandes comedores de semillas de cactus y de sus retoños famélicos, hasta el baby boom entre los pájaros de menor tamaño.
En casi treinta años de trabajo, los Grant han registrado algunos cambios extraordinarios en la estructura de la población y en su morfología, habiendo documentado extensamente significativas respuestas, tanto al nivel de población como al de especie, ante las evidentes presiones de selección debidas a un entorno con cambios tan drásticos. Registraron cambios en el tamaño de los picos y en su capacidad para romper semillas.
Las diferencias entre dimensiones del pico entre dos grupos de pinzones altamente especializados van desde ninguna (0 por cien) al 6 por ciento. Los nuevos trabajos sobre pájaros de Mayr y Diamond (2001), realizados en islas de Melanesia, evidencian una correlación inequívoca de las especies de pájaros con el aislamiento geográfico. Sin embargo, he aquí la cuestión crucial: ni en el Ecuador ni en Melanesia se ha registrado ningún caso de aparición de especiación, ninguno de los detalles que acompañan a la aparición de una nueva especie de pájaro.
Las diferencias en el tamaño del pico entre las seis especies distinguibles de pinzones terrestres oscilan alrededor del 15 por ciento. Ningún cambio de tal magnitud, relacionado con otros rasgos que permitieran hablar de una nueva especie de pinzón de las Galápagos, fue detectado ni por los Grant ni por ninguna otra persona. Así pues, el paradigma darwiniano funciona exactamente según lo previsto: los distintos rasgos (ya sea dentro de una misma especie o entre especies distintas) varían en prevalencia, de acuerdo con las exigencias el entorno. Obviamente, los genes que determinan esos rasgos cambian también. Sin embargo, no existe prueba alguna de que este proceso conduzca a la aparición de nuevas especies.
La ruta directa que nos permita remontar el curso de la especiación, sea ésta en las remotas Galápagos, en los cubículos de los drosophilósofos, o entre los montones de sedimentos de los paleontólogos, sigue por desvelar. Lo más próximo en ciencia a la observación y el registro de verdadera especiación en animales, consiste en el trabajo de Theodosius Dobzhansky sobre la mosca de la fruta Drosophila paulistorium. Pero incluso en este caso, no apareció ninguna especie nueva, sino tan sólo el aislamiento reproductivo, que tuvo lugar cuando una población plenamente fértil, que vivía entre temperaturas moderadas, se dividió en dos: una que prefirió vivir en el frío y otra que optó por el calor.
En nuestra opinión, la mejor prueba directa de especiación es también la menos conocida. Trata de seres asexuados y se oculta en la literatura profesional arcaica. Para comprobar directamente cómo una población de organismos da lugar a nuevas especies, hay que buscar entre los moradores del microcosmos. Para estudiar la especiación deberemos seguir el rastro de la simbiosis y bucear en la bibliografía sobre simbiogénesis. La inmensa mayoría de quienes escriben sobre evolución, tanto zoólogos como otros biólogos, tienden a ignorar la literatura existente acerca de la simbiosis y de la ecología de las comunidades de microbios u otros organismos.
La limitación intrínseca de la literatura darviniana fue analizada en 1999 por Douglas Caldwell, comenzando por el propio libro de Darwin de 1859. Entre los términos utilizados por Darwin y la cantidad de veces en que lo hace se encuentran: “batir” —diecisiete; “muerte” —dieciséis; “destruir (destruido, destrucción)” —setenta y siete; “exterminar (exterminación)” —cincuenta y ocho; “individuo” —doscientas noventa y ocho; “matar (muerto, matando)” —veintiuna; “perfecto (perfección)” —doscientas setenta y cuatro; “carrera (carreras)” —ciento treinta y dos; “seleccionar (selecciona, selección)” —quinientas cuarenta; “especies” —mil ochocientas tres. Como contraste, los siguientes términos están ausentes en El origen de las especies: “asociación, afiliación, cooperar, cooperación, colaborar, colaboración, comunidad, intervención, simbiosis”. Ciento cincuenta años más tarde persiste la costumbre de ignorar las asociaciones metabólicas y físicas entre organismos. En Análisis evolutivo (2001), un influyente libro de texto para universitarios escrito por Freeman y Herrón, los términos “combate”, “competición” y “conflicto” aparecen en al menos dieciocho páginas, mientras que “simbiosis” y “simbiogénesis” no se mencionan ni una sola vez en sus 700 páginas. Análisis eruditos sobre los «árboles de la vida», muchos de ellos calculados por ordenadores de alta velocidad, muestran masivamente ramas divergentes. Tan sólo en contadas ocasiones aparecen uniones de ramas para simbolizar alguna simbiogénesis. Tal vez esos gráficos de ordenador puedan ser muy impresionantes, pero no reflejan en absoluto ni la historia real de la vida sobre la Tierra, ni la evolución de las especies.
§. Herencia y salud
A lo largo de la mayor parte de la historia de la Humanidad, médicos, chamanes y herbolarios han venido practicando el arte de la curación en un estado de ignorancia bien educada. Si bien no existía nada parecido a la ciencia de la genética, los practicantes más astutos de este arte sabían que, en ciertas familias, prevalecían determinadas enfermedades. Por ejemplo, entre los hebreos de los tiempos bíblicos se consideraba imprudente circuncidar a un muchacho cuyo tío —exclusivamente por parte de madre— hubiera sangrado en exceso en su propia circuncisión. Para predecir la calvicie de una mujer o la enfermedad del «grito del gato» (Tay-Sachs) de su hijo, los galenos buscaban pistas en los antecedentes familiares de ambas ramas familiares. Durante más de cien años, la gran dinastía europea de los Habsburgo fue identificable por las mandíbulas prominentes de sus miembros.
Figura 2.1. El Árbol de la Vida.
Tanto las comadronas como los galenos meticulosos sufrían suficiente peste, fiebre puerperal, sífilis y demás formas de “virulencia” como para poder asimilar firmemente el concepto de “contagio”. Sin embargo, una filosofía coherente de la enfermedad contagiosa seguía mostrándose evasiva. Los hombres de medicina y los médicos forenses conocían bien la noción de enfermedad iatrogénica (causada por el galeno, según la raíz griega del término), convirtiendo la lealtad corporativa en un objetivo superior incluso al de no causar daño a sus pacientes. Los hospitales seguían siendo en muchos casos esa última parada que habían venido siendo a lo largo de la historia de Europa. Hasta que los trabajos de Louis Pasteur y de Robert Koch en el siglo XIX aportaran las pruebas de la identidad de los culpables de los contagios, ni los profesionales de la medicina ni el público en general sabían de la existencia de un microcosmos, de un universo de lo diminuto. Sólo después de que Pasteur descubriera las costumbres de las bacterias y los fermentos, y Koch desarrollara sus famosos postulados demostrativos de que «los gérmenes causan las enfermedades», comenzó a calar en nuestra herencia cultural la consciencia de la existencia de una presencia microbiana imperceptible a simple vista. No sería hasta la primera mitad de siglo XX, en parte gracias a una obra influyente escrita por Paul de Kruif, periodista que trabajaba en la Fundación Rockefeller de la ciudad de Nueva York, que la palabra “microbio” se convertiría en un término de uso corriente. La curiosidad científica está muy bien, pero son el temor y la codicia las que mueven a las masas. Al menos en los Estados Unidos, hasta que Cazadores de microbios de Kruif se convirtiera en el primer best-seller científico de la historia y los folletos gubernamentales comenzaran su campaña de consejos antisépticos, las madres jóvenes y los médicos octogenarios no comenzaron a tomarse realmente en serio que había que erradicar de nuestras vidas a las bacterias o “gérmenes”.
Al igual que “maleza” y “hongos”, “gérmenes” era un término indefinido pero claramente comprendido, que denotaba a formas minúsculas de vida, portadoras de enfermedades y por consiguiente indeseables. Considerados como «el agente enemigo», la conclusión lógica era —como sigue siendo aún para muchas mentalidades— que todo microbio debe ser derrotado. Ninguna diferenciación entre germen, virus, infección, microbio o bacteria era respetada. La recién nacida ciencia de la «microbiología» seguía sin saber nada acerca de las distintas fases del parásito de la malaria ( Plasmodio) o del intercambio nutricional entre la E. coli y su hábitat intestinal humano. Los médicos debían limitarse a clasificar por síntomas y prescribir un tratamiento seguro: humo, infusiones, aspirina, morfina, cocaína y extirpación, entre muchos otros “remedios”, constituían los recursos de sus maletines negros. A la desesperada, los pacientes eran sometidos a aplicación de ventosas, compresas, cataplasmas, cánticos o imposición de manos. Desde tiempos inmemoriales, el mejor antídoto consistía en la confianza dispensada a precio de oro por especialistas de autoridad, habitualmente hombres de gran tamaño y presencia imponente.
La teoría de la evolución estrena el siglo XXI en una posición muy parecida a la de la medicina de hace cien años. La búsqueda de nuevo conocimiento está inevitablemente embebida en la cultura. Los teóricos profesionales de la evolución tienden a ser abismalmente ignorantes en lo relativo a las tres ciencias —microbiología, paleontología y simbiosis— más relevantes para su trabajo. Pero la situación actual en relación con la evolución es incluso peor que su equivalente en la medicina del siglo XIX. La diferencia consiste en que, mientras que los datos de los que precisaban los galenos de finales del XVIII y principios del XIX no existían aún en sus días, las bases fundamentales para la ciencia de la evolución sí que existían a finales del siglo XX y principios del XXI, pero se prefiere ignorarlas sistemáticamente.
A pesar de que la mayor parte de los científicos parezca no querer darse cuenta de ello, la ciencia ha venido documentando la evolución en acción. La mayoría de la información significativa para la comprensión de la evolución sigue durmiendo entre la bibliografía oculta. Las noticias no llegan ni a los profesionales ni al gran público. Existe sin embargo un corpus de literatura fragmentado, detallado pero aún desorganizado, que documenta el modo en que se originan las especies y que recoge casos para el estudio de su evolución. Los fascinantes detalles acerca de cómo una especie conduce a otra, o sobre cómo se extingue para siempre, necesitan de portavoces. ¿Cómo aparece por primera vez la novedad compleja en seres vivos? ¿Cómo se extiende? ¿Cómo cambian con el paso del tiempo los cuerpos de plantas, animales y microbios? Se trata de una historia que aún no se ha escrito, porque la mayoría de sus anotaciones están registradas en los lenguajes esotéricos de las ciencias de la bioquímica y de la microbiología, entre otras. El relato del origen de las especies, junto con su relación con poblaciones aisladas, como explicara en su día Mayr (1942, 1982), ha permanecido en gran medida inaccesible, tanto a los lectores profanos como a los historiadores naturales y a los diseñadores de museos de ciencia. Incluso entre los biólogos y los geólogos que escriben acerca de la evolución, se da una gran pobreza de documentación y explicaciones sobre el historial de los orígenes de cualquier especie dada. En casos excepcionales, algunos botánicos llegan a establecer la relación entre diferencias en las células y los cloroplastos vegetales con la aparición de nuevas especies hortícolas o agrícolas. Sin embargo, habida cuenta de que estos especialistas no se ocupan en general de la evolución ni extienden sus explicaciones a los animales, las algas o las bacterias, sus descripciones tienden a quedar restringidas a un universo progresivamente disminuido de las «ciencias vegetales». Genetistas, ecólogos, microbiólogos, fisiólogos y demás gentes de laboratorio, junto con experimentadores, tienden a evitar el análisis de las implicaciones que de su trabajo se derivan para la evolución. La mayoría de ellos simplemente no tienen la menor idea acerca de cómo evolucionó la complejidad de la vida o, si la tienen, se abstienen de escribir sobre esta cuestión. En algunos casos, la herencia cultural que absorbieron con su formación académica les enseñó a desdeñar la historia evolutiva como una mera «especulación». Muchos de estos adeptos del «ala dura» ni siquiera consideran que los estudios sobre la evolución estén en el mismo nivel que las «ciencias duras» tales como la biología molecular o la bioquímica. Casi todos los científicos, por no decir su totalidad, prefieren habérselas con cuestiones a las que se pueda responder con pruebas directas. La reconstrucción de la historia de la vida, tarea central para los evolucionistas, se desprecia a menudo como poco más que un conjunto de especulaciones no demostrables. En los círculos científicos modernos, particularmente de los Estados Unidos, la filosofía y la historia son consideradas desde esta perspectiva como tradiciones cuestionables, como ocupaciones ociosas e inútiles. Los científicos en activo niegan incluso frecuentemente tener cualquier filosofía, asegurando que tal cosa interferiría con su trabajo. Sin embargo, la reconstrucción científica de la historia de la vida, práctica intrínsecamente filosófica repleta de inferencias y juicios, es esencial para poder dar respuesta a la pregunta fundamental de cómo se iniciaron las especies.
El aislamiento de los científicos contribuye asimismo a esta limitación. Incluso la comunicación entre campos relevantes para el estudio de la evolución, como pueden ser la paleontología y la biología molecular, se ve dificultada por diferencias terminológicas. Cualquier científico, por muy dotado que sea, no puede concentrarse más que en una sola cosa a la vez. En su quehacer cotidiano, el científico carece de cualquier posibilidad de integrar, en sus herméticos estudios, la miríada de descripciones idiosincrásicas de descubrimientos fundamentales sobre los organismos vivos. Es nuestro deseo conseguir abrir aquí algunas líneas de comunicación, con tan poca distorsión y sobresimplificación como resulte posible.
El fenómeno de la evolución ocurre sobre la totalidad de la faz de la Tierra, desde al menos catorce kilómetros bajo la superficie del mar, en los abismos oceánicos más profundos, hasta al menos ocho kilómetros por encima de su superficie, en las cimas de las montañas. La vida bacteriana ha sido detectada en fisuras húmedas de rocas graníticas, en profundidades de al menos tres kilómetros hacia el interior de la litosfera, así como en las chimeneas de agua caliente del fondo oceánico. La biosfera —el espacio donde la vida existe— presenta las dimensiones generales de una esfera hueca de unos veintitrés kilómetros de ancho lo cual, comparado con su radio de 6.371 kilómetros, corresponde tan sólo a un 0,0007 por ciento del volumen del planeta.
La vida comenzó probablemente hace algo más de 3.800 millones de años. El registro detallado de la evolución, conservado en las rocas tanto en forma de fósiles como de compuestos de carbono de cadena corta y larga extraíbles, desborda a quienes pretendan estudiarlo. La vida celular refleja su historial evolutivo. Sin embargo, a pesar de la diversidad de indicios a través de los que resulta posible reconstruir la evolución de la vida, la mayor parte de los autoproclamados biólogos evolutivos desdeñan la biología celular, la microbiología e incluso el registro geológico de las rocas. Muchos de ellos se encuentran tan ocupados con los animales terrestres que siguen creyendo que no existe registro alguno de la evolución que pueda remontarse más allá de los últimos 541 millones de años. El crucial estudio científico de los eones Arcaico y Paleozoico, la geografía global del metabolismo y sus detalles, junto con la química, contienen sin duda la respuesta al enigma darwiniano, a pesar de lo cual estos vastos campos de búsqueda se ven formalmente excluidos de toda consideración.
En la medida en que la bioquímica, la microbiología y la microscopía electrónica han ido subdividiéndose, los conceptos evolutivos se han vuelto necesariamente cada vez más multidimensionales e interconectados. La inmensa expansión de la bibliografía sobre biología molecular, particularmente en lo concerniente a los datos sobre las secuencias de las proteínas y los ácidos nucleídos —moléculas siempre presentes en los seres vivos—, ha aportado nuevas y sustanciales profundizaciones a la visión de Darwin. Los detalles de la evolución molecular apoyan la generalización que postula que tanto las secuencias de aminoácidos en las proteínas como las secuencias nucleótidas en el ADN y en el ARN están relacionadas por un pasado común. Sin embargo, el abstracto gen dominante «A» que muta hasta convertirse en su alterno recesivo (alelo), que propone la teoría neodarwinista, constituye una vasta e incluso inútil sobregeneralización del verdadero cambio genético en las poblaciones. En lugar de las idealizadas formalidades de la «síntesis moderna» darwiniana, los principios organizadores necesarios para la comprensión de la vida requieren nuevos conocimientos en química y en metabolismo. Desde que Darwin y sus seguidores publicaran sus estudios al respecto, la profundización en el conocimiento sobre el funcionamiento de las células ha permitido clarificar las modalidades de evolución. Los resultados de la nueva ciencia, tanto en el laboratorio como sobre el terreno, contradicen, rebasan o marginan el formalismo del darwinismo, excepto en lo relativo a las variaciones en el seno de poblaciones de mamíferos y otros organismos de reproducción sexual. Los mamíferos constituyen, probablemente, tan sólo una diezmillonésima parte de las especies hoy en vida.
Los agentes del cambio evolutivo tienden a ser organismos plenamente vivos —microbios y sus relaciones ecológicas— y no tan sólo las mutaciones aleatorias que suceden en su interior. La formación y la diversificación de toda nueva especie es la manifestación externa de las acciones de formas de vida subdivisibles: los microbios más pequeños, las bacterias y sus descendientes de mayor tamaño, microbios más grandes, protistas y hongos, junto con sus legados intracelulares, orgánulos tales como las mitocondrias y los centriolos. La evolución surge del hecho de que estos pequeños organismos vivos y su progenie tienden a rebasar sus perímetros. Esos seres invisibles que diezman nuestras poblaciones con enfermedades virulentas, pero que también aportan al suelo el nitrógeno necesario para las plantas de las que nos alimentamos, juegan el papel principal en la génesis de las nuevas especies.
§. Células, microbios, propágulos y entidades autopoiésicas
La unidad mínima de vida es la célula. Todos los microbios se componen de células. Pueden ser clasificados inequívocamente en bacterias (sin núcleo) o protistas y hongos, ambos formados por células con núcleo. Las células de menor tamaño y menos complejidad, como por ejemplo la Mycoplasma geniticulum, contienen aproximadamente 500 genes.
Algunas personas, y de hecho algunos autores, presentan a los virus como la forma menor de vida. Sin embargo, los virus no están vivos e incluso son, en principio, demasiado pequeños como para ser considerados unidades de vida. Carecen de los medios necesarios para la producción de sus propios genes y proteínas. Los virus se comportan como elementos químicos hasta penetrar en las células vivas adecuadas, donde se aprovechan del entorno celular para reproducirse. Las células, en cambio, cuando disponen de acceso a materia y energía utilizables y se bañan en el medio acuoso adecuado, crecen —habitualmente hasta doblar su tamaño— y se reproducen, generalmente por división. Las células exhiben metabolismo, es decir, reacciones químicas interconectadas que mantienen sus procesos. Como los virus, los genes son incapaces por sí mismos de producir material celular que, en su mayor parte, consiste en proteínas. Tanto los virus como los genes precisan de células vivas intactas para su crecimiento, su reproducción y su propagación. Mientras que las células más elementales pueden tener 500 genes o menos (y por consiguiente producir 500 proteínas diferentes o menos), la mayor parte de las células bacterianas contienen entre 2.000 y 5.000 genes. Otras, más complejas y de mayor tamaño, como las células bacterianas respiradoras de oxígeno, pueden llegar casi a los 10.000 genes. Tales bacterias se aproximan por consiguiente al número de genes típico de las células de los organismos nucleados. Las células nucleadas, componentes básicos de la totalidad de los grandes organismos (hongos, animales, plantas y sus antepasados protoctistas), contienen habitualmente de 10.000 a 100.000 genes. Las células humanas suelen tener entre 30.000 y 60.000 genes, mientras que la Saccharomyces cerevisiae, la levadura que solemos emplear para la confección de pan y cerveza, tiene tan sólo unos 8.000.
El término “autopoyesis” (literalmente construirse a sí mismo) se refiere a la química de automantenimiento característica de las células vivas. Ningún objeto material menos complejo que una célula puede mantenerse a sí mismo y a su propio perímetro con una identidad que le distinga del resto de la Naturaleza. Las entidades vivas autopoiésicas mantienen activamente su forma e incluso la cambian a menudo (se «desarrollan»), pero siempre a través del flujo de materia y energía. En cualquier organismo, en cualquier entidad autopoyésica, es posible identificar una fuente específica de energía sustentante (luz visible, oxidación del metano o del sulfuro), junto con otra de carbono (azúcar, proteína, dióxido de carbono), nitrógeno u otros elementos químicos necesarios.
A temperaturas próximas a los cero grados, donde el movimiento de las moléculas se detiene, el metabolismo —la manifestación de autopoyesis— cesa también. Sin embargo, la estructura material de numerosas células vivas conserva la capacidad de reanudar el metabolismo cuando la temperatura asciende de nuevo. Las esporas bacterianas y los embriones cuidadosamente congelados no pierden su organización intrínseca con la congelación, por lo que al recibir calor de nuevo son plenamente capaces de reanudar las transformaciones metabólicas que aseguran su automantenimiento. Podemos, pues, afirmar que, aunque la vida florezca plenamente únicamente como resultado del flujo de materia y energía, puede también permanecer durmiente en la organización de la materia viva.
Los microbios son verdaderos especialistas en pasarles su ADN a otros en forma de genes enteramente funcionales. Estas artimañas sustentan la historia de la evolución darwiniana. Los microbios abandonados a su suerte, en condiciones de estrés y privaciones, tienden a unirse a otras formas de vida. Algunas de estas asociaciones duran una estación, o incluso menos, pero ocasionalmente los microbios se funden permanentemente con formas de vida superiores. Lamarck estaba en lo cierto: los rasgos adquiridos pueden serlo no como tales, sino como genomas. Sin embargo, tanto Lamarck como Darwin, con su idea de pangénesis, se equivocaron al sugerir que la adquisición pudiera ser el destino de toda característica. El único «carácter» o rasgo que puede ser transmitido hacia abajo (por herencia vertical) o adquirido (por herencia horizontal), y luego propagado de generación en generación, es el que esté codificado en genes. Ello significa que los «caracteres» susceptibles de ser propagados en el entorno, por adquisición o herencia de una generación a la siguiente, tenderán a ser genes completos dentro de genomas. Los genomas son sistemas genéticos enteros, activos únicamente cuando residen en el interior de la célula. Los genomas ajenos pueden ser —y de hecho lo son frecuentemente— heredados por otros. Los seres vivos compuestos por células pueden transmitir —y así lo hacen— conjuntos de genes a otros seres vivos. ¿Por qué han tenido tanto éxito los ingenieros genéticos en la transmisión de genes ajenos a vegetales comestibles y animales domésticos? Porque la Naturaleza lleva eones refocilándose en esta clase de artimañas de intercambio de genes y de genomas. El verdadero ingeniero genético es el microbio; los científicos y los técnicos son meros intermediarios.
Entre los protistas encontramos los mejores ejemplos de que las nuevas especies se forman gracias a la herencia de microbios adquiridos. Sabemos que el proceso de herencia de un conjunto de genes ajenos no es continuo ni premeditado. La Naturaleza no tiende a ser previsora, sino más bien oportunista. Sin embargo, la adquisición de genomas ha dejado en numerosas ocasiones tantas pruebas de su paso, que resulta factible seguir sus enrevesadas huellas. La mayoría de absorciones, de adquisiciones, de fusiones y de uniones genéticas tienen lugar en condiciones de dificultades medioambientales. La complejidad y la capacidad de respuesta de la vida, manifestadas particularmente en la aparición de nuevas especies a partir de fusiones entre antepasados distintos, tan sólo pueden ser comprendidas a la luz de una peculiar historia de la evolución. En cada caso es necesario formular las mismas preguntas: ¿Cuándo, dónde y en qué poblaciones de organismos evolucionó la nueva especie? ¿Con qué frecuencia y en qué grupos de organismos constituye la adquisición de genomas la modalidad principal de especiación? Ésta es la clase de cuestiones de investigación que aspiramos a suscitar.
La saga evolutiva presente, legítimamente vulnerable a la crítica tanto desde dentro como desde fuera de la ciencia, se vuelve incomprensible cuando se elimina del escenario a los microbios. Del mismo modo que la química resultaría incomprensible sin la mención del átomo, la dialéctica evolutiva carece en gran medida de sentido sin la comprensión previa del microcosmos.
§. Termodinámica, «propósito» y «progreso» evolutivo
La aceleración del cambio se nos lleva por delante. Una semana uno de los autores (Lynn Margulis) pasaba con su bicicleta, camino del campus como de costumbre, ante una casa pintada de amarillo, repleta de estudiantes. A la semana siguiente nada obstruía la vista desde la calle al patio de la escuela del barrio. En el horizonte, las montañas dibujaban su perfil en la distancia. La casa amarilla había desaparecido. No quedaba ni rastro de su existencia, a excepción tal vez de las huellas de su implantación sobre el suelo removido. Estos cambios espectaculares en nuestro entorno más inmediato se han vuelto corrientes. Burguer Kings, Toys “R” Us, Wendy’s, MacDonalds y sucursales bancarias brotan por doquier en nuestras ciudades y en nuestro pueblos. Las tiendas de la abuela, los campos de trigo y los robledales desaparecen como monedas en la arena. Los indios americanos prosperan si se unen para formar asociaciones de juego, los estudiantes graduados reciben subvenciones si dejan de estudiar las costumbres de los castores en sus madrigueras para ponerse a investigar sobre el genoma humano. ¿Por qué será que las fuerzas del cambio parecen prevalecer siempre sobre los tranquilos e inmutables hábitos del pasado? ¿Por qué parece la evolución de la vida acelerar a medida que avanzamos desde el presente hacia el futuro? ¿Es la evolución un mero cambio evolutivo? ¿No parece más bien que el propio proceso evolutivo, el origen y la diversificación de la vida a partir de unos antepasados comunes, estuviera dirigido? Cuando preguntamos a los biólogos evolutivos y a otros científicos si la evolución de la vida va en alguna dirección, lo niegan rotundamente. Sin embargo, nuestra experiencia cotidiana nos sugiere que nuestro entorno social se vuelve cada día más complejo. Nuestro entorno natural acuático y verde parece encogerse cada vez más, cediendo paso a un paisaje de acero y hormigón: luces de neón, señales de tráfico y demás aspectos de la urbanización reemplazan a bosques y corrientes de agua a una velocidad siempre creciente. Las personas desplazan de su hábitat a zorros y antílopes; donde antes medraban oropéndolas y pájaros carpinteros proliferan ahora palomas y gorriones. Las herramientas digitales substituyen a los ingenios mecánicos a una velocidad alarmante. La evolución de la vida sí que parece tener una dirección. Las peculiaridades de la vida y de las tecnologías humanas parecen expandirse a un ritmo acelerado de cambio, a medida que avanzamos del presente hacia el futuro.
Aquí puede resultamos de utilidad la termodinámica, una rama de la física matemática. Los termodinámicos describen las máquinas y la química en compartimentos estancos. Tradicionalmente han venido trabajando con la energía, la temperatura y el calor y, como científicos de una disciplina física que son, han venido evitando deliberadamente a los seres húmedos y sinuosos. Pero nos encontramos en el umbral de un gran cambio: la unión de la ciencia exacta de la termodinámica con otras ciencias tales como la biología, la meteorología y el cambio climático. La Naturaleza sigue su curso y los científicos aprenden, después de numerosos fracasos y de seguir a tantas estrellas falsas, a describir adecuadamente a la Naturaleza en términos matemáticos y químicos. Parece probable que la nueva termodinámica de la vida vaya a ejercer una influencia perdurable sobre el pensamiento evolutivo, en la medida en que conecta a la materia viva con la materia inerte de forma comprensible.
Todos sabemos que «la naturaleza aborrece el vacío» y que, por consiguiente, el aire se precipita a llenar cualquier vacuidad que produzcamos con una bomba. Una lata vaciada de aire se colapsa inmediatamente. También sabemos que la gravedad atrae a los cuerpos hacia el centro de la Tierra y que, por consiguiente, cualquier cuerpo arrojado al vacío caerá hacia abajo. Tomamos esa caída predecible de la materia al suelo como una ley más de la Naturaleza.
La nueva termodinámica une ambos conceptos en la frase «la naturaleza aborrece el gradiente». A medida que aprendemos a comprender esa novedosa idea fundamental se abre ante nosotros una perspectiva radicalmente nueva: la vida se comporta según las leyes de la Naturaleza, exactamente del mismo modo en que las moléculas de aire se apresuran a llenar el vacío, o en que los cuerpos caen irremisiblemente al suelo. Así pues, ¿en qué consiste esta nueva termodinámica y cómo funciona?
En The Way of the Cell (2001) Franklin Harold escribe: «Quienes se plantean un vínculo fundamental entre la flecha de disipación de la energía de la termodinámica y la flecha de la Tierra reverdecida de la biología son hoy una pequeña minoría… Pero si su visión es acertada, revelaría una profunda continuidad entre la física y la biología, el manantial de vida definitivo». Nos encontramos entre quienes opinan que esta visión es certera. La evolución es la ciencia de la conexión, y ésta no termina con los vínculos entre humanos y simios, o entre éstos y otros animales, o entre otros animales y microbios: lo vivo y lo no vivo están también relacionados de formas fundamentales. Hemos visto que la organización de la vida es material, pero la energía también se organiza.
Eric D. Schneider, geólogo de la Universidad de Columbia que sirvió en los años 70 en la U. S. Environmental Protection Agency [6], es uno de los principales visionarios de la nueva termodinámica. Disgustado con el primitivo nivel de ciencia practicado en la E. P. A., donde se experimentaba con peces colocados en tarros de pepinillos etiquetados y sometidos a niveles variables de toxinas para determinar el «grado apropiado de contaminación», Schneider, que consideraba que la protección del medio ambiente debía estar basada en una adecuada descripción de los fenómenos ecológicos, dedicó décadas de su vida a investigar de qué modo lo no vivo está relacionado con lo vivo. Partiendo de los trabajos de científicos como L. Boltzmann, V. I. Vemadsky, I. Prigogine, J. E. Lovelock y H. Morowitz, sus descubrimientos son simples y a la vez reveladores: la vida se encuentra en una clase de sistemas que se organizan en respuesta a un gradiente. El gradiente se define como una diferencia a lo largo de una distancia.
Todos estos sistemas, entre los que se cuenta la vida, producen características sorprendentemente complejas. Los gradientes de temperatura provocan fluidos altamente organizados, corrientes octagonales conocidas como células de Bénard. Los gradientes de presión atmosférica producen tomados y patrones climáticos globales, que desaparecen cuando se nivelan las presiones barométricas. Los gradientes químicos dan lugar a complejos “relojes” químicos[7]. Se hace necesario señalar aquí que no tan sólo es la vida coherente con la segunda ley de la termodinámica —la entropía crece en los sistemas aislados—, sino que los sistemas complejos (desde los tomados que nivelan gradientes barométricos hasta los organismos que reducen gradientes de energía solar o química) tienden a reducir los gradientes en su entorno de forma más rápida y eficaz que si no existiesen. Un gradiente de presión atmosférica, por ejemplo, tardaría más en alcanzar el estado de caos aleatorio sin el complejo sistema cíclico del tomado, cuya función consiste precisamente en alcanzar ese fin natural.
Los gradientes originales descritos por la ciencia se referían a los motores a vapor. Las diferencias entre calor y frío, que el ciudadano corriente hacía tiempo que se había dado cuenta de que tendían a igualarse por su propia cuenta hasta convertirse en tibio, podían ser conveniente transformadas en energía por la máquina. Las matemáticas necesarias para convertir el calor en trabajo constituyeron la base de la ciencia de la termodinámica. El término tiene sus raíces griegas en “calor” y “movimiento”. La termodinámica constituye el estudio sobre cómo los sistemas manejan y transforman la energía. La termodinámica clásica se ocupaba de sistemas aislados y más o menos cerrados, que acababan por perder su funcionalidad y cuyas moléculas constituyentes terminaban por disponerse aleatoriamente, hasta el punto de no poder seguir funcionando.
Pero los seres vivos no son máquinas de vapor ni sistemas mecánicos cerrados. Somos sistemas abiertos, organizados a partir del flujo de materia y energía que circula incesantemente a través de nosotros. Por consiguiente nosotros (la vida, en un sentido genético amplio) no estamos destinados a revertir a un estado final aleatorio, medido ya sea en sistemas químicos aislados o en máquinas abandonadas a su suerte. La diferencia entre los sistemas vivos abiertos y los sistemas artificialmente cerrados de la termodinámica clásica determinó que esta ciencia de la energía fuera considerada en un tiempo como irrelevante para la vida. Peor aún, en los siglos XIX y XX fueron numerosos quienes entendieron ingenuamente que la teoría darwiniana de la evolución por medio de selección natural significaba que la termodinámica contradecía el fenómeno de la vida. Los sistemas termodinámicos se tomaban aleatorios, desorganizados y menos complejos con el paso del tiempo, mientras que los sistemas vivos incrementaban visiblemente sus niveles de organización y complejidad. La nueva termodinámica ha venido a resolver la aparente contradicción entre la tendencia a la desorganización de la materia inerte y la tendencia a la organización de la materia viva. La vida no existe en un vacío, sino que ocurre en la diferencia muy real que media entre una radiación solar de 5.800 Kelvin y las temperaturas de 2,7 Kelvin del espacio exterior. Ése es el gradiente del que se nutre la complejidad de la vida. Como en el caso de las células químicas de Bénard, que se estabilizan desafiando las tendencias estadísticas a aproximarse al equilibrio manifestando el improbable gradiente que las rodea, como los remolinos meteorológicos que mantienen su estructura «para» liberar una diferencia entre presiones atmosféricas, la vida es también un sistema de reducción de gradientes. Estas ideas de la termodinámica relacionan lo vivo con lo no vivo, del mismo modo que la selección natural de Darwin nos relaciona con los grandes simios y, en último extremo, con los microbios. El poder de la termodinámica para explicar las características generales de la vida no es en absoluto cuestión banal, bien al contrario: es parte integrante del futuro de la ciencia.
Así pues, la vida no contradice la bien conocida segunda ley de la termodinámica, ni hace tampoco falta ninguna ley nueva, una vez que nos hemos percatado de que el estudio del comportamiento complejo a cargo de la termodinámica clásica estaba limitado por su uso restrictivo de sistemas cerrados y aislados. Cuando el ámbito de la termodinámica clásica es extendido al cosmos abierto, la complejificación se hace comprensible. En lugar de tratar de acuñar precipitadamente una «cuarta ley» de aplicación a la vida, como proponía Stuart Kauffman en 1999 en su libro Investigaciones, podemos aplicar tranquilamente la visión general de la termodinámica a la totalidad de la materia. La tecnología, sea ésta humana, del ave del Paraíso o de la bacteria fijadora de nitrógeno, se convierte entonces en una extensión para los sistemas abiertos de la segunda ley de la termodinámica. Esta segunda ley, que describe un inexorable incremento en el parámetro conocido como entropía —formulado inicialmente como «calor dividido por temperatura» y más adelante convertido en la tendencia estadística a las disposiciones más probables de materia y energía a lo largo del tiempo— no contradice en absoluto la tendencia local de la vida a incrementar el orden, sino todo lo contrario. Los creacionistas se equivocan. El origen de la vida, su organización celular, su expansión y la evolución de biodiversidad creciente, son enteramente coherentes con el nuevo pensamiento de la termodinámica [8]. Las asombrosas —aunque no milagrosas— disposiciones de la materia viva proporcionan un ejemplo más, junto con los de los huracanes circulares y los relojes químicos, sobre cómo construye la Naturaleza estructuras destinadas a reducir gradientes. La mayoría de evolucionistas argumentan que la vida se originó en la Tierra, tal vez por causa de un determinado gradiente químico entre compuestos ricos en hidrógeno y otras substancias ricas en carbono y oxígeno. Se trata de combinaciones combustibles. O tal vez la vida tuvo su origen en la luz, cuando los fotones solares de alta energía entraron en el ciclo de los precursores del metabolismo, antes de que éste evolucionara hacia su estado moderno[9]. La vida desarrolló la identidad de la célula. La materia viva en forma de células y organismos constituidos por ellas comenzó a reducir los gradientes químicos y energéticos de un planeta aún adolescente y turbulento. De hecho, la tendencia a organizarse localmente «para» liberarse de las anomalías estadísticas encamadas por los gradientes es profunda y determina la organización de la vida a todas las escalas. La eficacia superior de los ecosistemas complejos para reducir gradientes es mensurable y ha sido medida, por ejemplo, por termómetros aéreos que demuestran la capacidad de las selvas tropicales para refrescarse, superior a la de los prados y los desiertos, reduciendo así el gradiente solar electromagnético. No se trata, pues, de una teoría vaga y abstracta, sino de una hipótesis comprobada: tanto en mediciones por aeroplanos a baja altura como por satélites, los ecosistemas son más frescos cuanto más maduros y biodiversos son. Los ecosistemas se inician a partir de especies colonizadoras de crecimiento rápido y, al principio, su eficacia como reductores de gradientes es escasa. Sin embargo, a medida que van madurando, sus ciclos de materia y energía van aumentando el ámbito de acción. Entran en el sistema nuevos organismos que establecen nuevos hábitats. El crecimiento se enlentece pero, en conjunto, el ecosistema integrado se vuelve más eficaz en la reducción del gradiente entre el Sol y el espacio. El hecho de que los ecosistemas tropicales maduros permanezcan frescos demuestra el poder del sistema para reducir gradientes [10].
No es tan sólo el desarrollo de los ecosistemas el que viene informado por el mandato termodinámico, por esa «ley de la Naturaleza» que ordena reducir gradientes, sino también la dirección de la evolución como un todo. Tanto en los ecosistemas como en la evolución y en las economías, detectamos un aumento de la complejidad, por ejemplo, en el número total de extensión de especies sobre un volumen mayor, en la creciente diferenciación de la «división del trabajo», en redes informáticas cada vez más desarrolladas y complejas, o en la creciente integración funcional de flujos de materia y energía en el entorno urbano tales como el gas, el agua, la electricidad o la red de alcantarillado. El aumento de las capacidades de los organismos para adaptarse a recursos actualmente en disminución constituye un ejemplo adicional. A pesar de que resulte difícil explicar por qué el universo está tan organizado, la comprobada expansión del espacio desde el Big Bang sigue proporcionando un «sumidero» (un lugar) hacia el que las estrellas pueden irradiar. La expansión del cosmos produce de este modo un gradiente cósmico, crisol a su vez de todos los demás gradientes. Por lo que se refiere al futuro predecible como mínimo, la geometría de la expansión del universo seguirá generando posibilidades para la destrucción funcionalmente creativa de gradientes, por ejemplo en relación con el espacio y con el gradiente electromagnético de las estrellas. Una vez que comprendemos esta organización, la vida ya no se nos aparece como un milagro, sino más bien como un sistema cíclico más, con una dilatada historia, cuya existencia queda explicada por su eficacia en la reducción de gradientes, superior a la de los sistemas no vivos a los que reemplazó.
Esta nueva termodinámica —en ocasiones llamada también homeodinámica— nos permite comenzar a atisbar el camino desde la materia (colapso de gradiente) a la mente (percepción de gradiente); de la «auto»-organización energética a la informacional. Escribimos «auto» entre comillas porque, en realidad, los sistemas complejos son precisamente «otro»-organizados, nunca organizados por sí mismos. La tendencia de los sistemas a organizarse no proviene de sus componentes internos, sino de los gradientes presentes en su entorno inmediato. La estructura informacional de la vida, transportada en el ADN, se ha vuelto auto sustentadora por conducto de la reproducción. Sin embargo, la memoria existe igualmente en sistemas completamente no vivos, tales como los vórtices organizados por gradientes de presión atmosférica rotativos. Estos sistemas saltan a nuevos estados y nuevos valores que dependen de su historia. Nos parece pues que, sin necesidad de invocar ninguna clase de vitalismo, misticismo o espiritualidad, podemos reconocer en nosotros mismos cierto «propósito», fruto de la tendencia termodinámica a avanzar hacia el equilibrio. Los sistemas complejos —incluyendo a la vida— tienden a emerger para aproximar hacia el equilibrio a sus entornos ricos en gradientes.
A los ojos de muchos cristianos, la revolución darwiniana dejó a la Naturaleza despojada de propósito, al menos sobre el papel. Los darwinianos, enfrentados ante un Creador personal como única fuente concebible de propósito, se apresuraron a mostrarse de acuerdo. Pero el propósito físico es mucho más sutil que todo eso. Desde el ventajoso punto de vista de la termodinámica, el propósito tiene un aspecto físico. El propósito no es más uniforme que la memoria —que se manifiesta genéticamente en los cuerpos y neuronalmente en los cerebros, e incluso magnéticamente en las máquinas— y al igual que ella, el propósito —con su orientación hacia el futuro— tiene una génesis termodinámica.
La vida es termodinámica. El remolino continuo que desciende más abajo de las cataratas del Niágara tiene un nombre: «remolino». Les ponemos nombres a las cosas —como a las especies o a los huracanes— que mantienen una identidad, al menos durante cierto tiempo. La formación de identidades estables contribuye al proceso termodinámico de reducción de gradientes. Los miembros altamente heredables de una especie proporcionan, del mismo modo que otros agentes termodinámicos cíclicos y complejos, vehículos estables de degradación. Los seres cíclicos de la vida sobreviven para reducir los gradientes energéticos y materiales que les mantienen en marcha. Ambicionan esos gradientes y se nutren de ellos, al menos durante el tiempo suficiente como para reproducirse. En la medida en que la selección natural filtra a muchos para conservar a unos pocos, esos pocos utilizan de forma aún más eficiente la energía disponible en el entorno para reducir «a propósito» sus gradientes. La cuestión crucial radica en que los «seres», tanto vivos como no vivos, devienen para reducir gradientes de forma natural. El ser reproductor de la biología es un ciclo de orden elevado cuyos antecedentes pueden ser inferidos a partir de los ciclos del mundo no vivo. La replicación nucleótida y la reproducción celular no surgen de la nada, sino que nacen en un universo energético a partir de las tendencias termodinámicas inherentes en la Naturaleza.
Capítulo 3
Individualidad relativa
Individualidad dentro de la comunidadNos resulta fácil reconocer a los individuos. Denominamos población a un grupo de individuos de la misma especie, en un mismo lugar al mismo tiempo. Junto a otras formas de vida, por ejemplo plantas y animales susceptibles de servir como alimento, decimos que distintas poblaciones al mismo tiempo y en un mismo lugar forman comunidades. En determinadas ocasiones, puede que los miembros dominantes de esas comunidades tengan menos de un milímetro en su mayor dimensión. Cuando esto sucede, decimos que se trata de una comunidad microbiana. Ninguna manifestación de vida sobre la Tierra consiste —al menos no durante cierto tiempo— en miembros aislados de una misma población, como adolescentes encarcelados o como filas de plantas de maíz.
La fuerza del número
Las especies desconstruidas: la ardisia de miehe y la euplotidium de rosati
El secreto del borde de la hoja de miehe confirmado
Ciliados defensivos o la revelación de rosati
Comunidades y la lógica de los ecosistemas
§. Individualidad dentro de la comunidad
Las comunidades son grupos naturales, que podemos reconocer en el mundo natural fácilmente, sin necesidad de una formación específica: comunidades arborícolas, comunidades acuáticas, comunidades costeras, comunidades del acantilado o comunidades del bosque. Como regla general, que empleamos de forma semiconsciente, decimos que los individuos que parecen tener todo lo importante en común, que parecen ser «lo mismo» en los principales aspectos de su vida, pertenecen a una misma especie.
En el siglo XVII, Linneo, algunos naturalistas y otras personas interesadas en la historia natural, comenzaron a poner nombres a las especies de todo el mundo y a documentarlas. A medida que los europeos exploraban los trópicos de América, África y el Sudeste de Asia, comenzó a fluir un verdadero torrente de documentos y publicaciones al respecto. La tarea linneana de documentar y nombrar adecuadamente a todos los organismos nunca ha sido completada. Incluso en nuestros días está teniendo lugar un esfuerzo para animar a las corporaciones globales a que destinen parte de sus beneficios a la realización de un «Inventario Total de las Especies». Con la utilización internacional de ordenadores de alta velocidad, de tecnologías por satélite y de transmisión en tiempo real de grandes cantidades de datos, el líder de este proyecto, el naturalista Peter Warshall, argumenta que deberíamos estar en condiciones de registrar la totalidad de la biodiversidad del planeta antes de que la destruyamos definitivamente. Como una muerte en la familia, la extinción de cualquier especie constituye una pérdida irreparable.
Aun cuando el «Conjunto de Datos sobre Todas las Especies del Planeta» dista mucho de estar completo, de su estado actual comienzan a surgir algunas características notables. Se estima que hay hoy en vida entre diez y treinta millones de especies distintas de organismos, mientras que tan sólo han quedado documentadas en la bibliografía profesional menos de dos millones. El número de especies de organismos extinguidos, registradas en la literatura paleontológica basada en las pruebas fósiles de su existencia pretérita, se limita a unas 150.000. La mayoría de los científicos está de acuerdo en que el 99 por ciento de las especies que en algún momento han existido, están hoy extintas. De ello se deduce que el número total de especies que han poblado el planeta Tierra desde que la vida se iniciara sobre él, hace unos 4.000 millones de años, ascendería a 200 billones. Nadie que conozcamos es capaz de distinguir más de unos pocos millares de especies y, en realidad, no conocemos prácticamente a nadie que pueda reconocer a ese número, ni siquiera con la ayuda de los oportunos textos especializados. Sobre el terreno, particularmente en los flagrantemente biodiversos trópicos del nuevo mundo, la mayor parte de nosotros nos sentimos impotentes. Incluso Mayer Rodríguez, consumado guía ecuatoriano de la rivera tiputini del Napo, afluente del Amazonas, no consigue identificar más allá de 500 especies. Y nadie conoce la región mejor que él.
Todos los organismos conocidos pueden ser inequívocamente catalogados en uno de dos grandes grupos, diferenciados por la clase de células que los constituyen. El primero de ambos grupos, presumiblemente también el de más temprana aparición, es, con mucho, el más diverso (desde el punto de vista de modalidades metabólicas) y también el más esencial para el entorno sobre la superficie del planeta. Se trata de las bacterias, todas las cuales están compuestas por células que carecen de núcleo, denominadas procariotas. Las formas de vida que nos resultan familiares (plantas y animales), así como los dos grupos de seres vivos más pequeños pero visibles (hongos y protoctistas) pertenecen al segundo grupo, de aparición más reciente. Estas formas de vida mayores se conocen con el nombre de eucariotas, organismos verdaderamente nucleados, es decir, cuyas células tienen núcleo.

Para las cantidades de especies vivientes de estos reinos disponemos tan sólo de toscas estimaciones, que podrían ser tremendamente erróneas. Dominan los animales, probablemente porque la gente sabe distinguir bien a los escarabajos. Se cree que pueden existir más de 10 millones, tal vez 30. Se sugiere que pueden estar medrando en los bosques y las aguas de este mundo unas 50.000 plantas, unos 100.000 hongos y unos 250.000 protoctistas. Por lo que se refiere a las bacterias, aunque miles de ellas han sido nombradas como especies y no cabe duda de que pueden distinguirse otros tantos millares, no les es de aplicación el concepto de especie. A pesar de que puedan ser agrupadas según sus características comunes, estos grupos cambian con tanta rapidez que nunca resulta posible reconocerlas como en el caso de las especies eucariotas. Las bacterias intercambian genes libremente. Todas ellas pueden reproducirse por sí solas, pudiendo así descender en cualquier momento de un solo progenitor. La intervención del sexo, la formación de una nueva bacteria con genes de más de una fuente única, constituye una cuestión unidireccional. Los genes pasan de un individuo donante a otro receptor… pero los donantes pueden convertirse en recipientes y viceversa en cuestión de minutos. Es más, el intercambio de genes es completamente opcional. Si una bacteria logra sobrevivir y desarrollarse en las condiciones en las que se encuentra, puede prescindir en cualquier momento del sexo. De hecho, las bacterias están dispuestas y capacitadas para «practicar el sexo» con las moléculas desnudas de ADN que absorben del medio acuoso en que se bañan.
La vida se originó con las bacterias y, por consiguiente, es lícito afirmar que el origen de la vida concurre con el origen de las bacterias. Sin embargo, estamos de acuerdo con el profesor Sorin Sonea y su colega Lucien Mathieu, de la Universidad de Montreal, en que las bacterias no tienen especies en absoluto, o en que —lo que equivale a lo mismo— todas ellas en conjunto constituyen una especie única y cosmopolita. La especiación es una propiedad exclusiva de los organismos nucleados. Se inició con los primeros protoctistas, mucho después de que las bacterias hubieran desarrollado la mayoría de los rasgos metabólicos importantes, desplegados por la vida sobre la Tierra. Por consiguiente, el origen de las especies no fue en sí mismo concurrente con el origen de la vida, sino que ocurrió mucho más tarde, en el Eón Proterozoico. La individualidad no proviene exclusivamente de la diversificación y de la ramificación de la evolución, como desearían hacemos creer los neodarwinistas, sino que proviene con igual frecuencia, sino incluso mucho más a menudo, de la integración y diferenciación de seres fusionados, otrora independientes, pero con el paso del tiempo individualizados y seleccionados como un todo único. Se trata simplemente de una forma más de formular el argumento central de nuestra tesis: la especiación por medio de (y en la resaca de) la simbiogénesis.
La fuerza creativa de la simbiogénesis produjo células eucariotas a partir de bacterias. Por consiguiente, todos los organismos superiores —protoctistas, hongos, animales y plantas— se originaron simbiogenéticamente. Sin embargo, la creación de novedad por medio de la simbiosis no acabó con la evolución de las primeras células nucleadas, sino que la simbiosis sigue presente por doquier
Son numerosos los ejemplos de evolución por simbiosis que asombran por su belleza. Los arrecifes coralinos del Pacífico, tales como la Gran Barrera de Australia, representan asociaciones entre corales modernos (escleractinios) y dinomastigotes tales como el Gymnodinium microadriaticum. Los líquenes de Nueva Inglaterra, las plantas hormigueras de Nueva Guinea, e incluso las vacas lecheras, sirven de ejemplo del poder de las fusiones entre seres vivos. Miembros de distintas especies —y en el caso de los corales y las vacas incluso de reinos distintos— forman, bajo condiciones de estrés identificables, comunidades íntimamente ligadas, que acaban convirtiéndose por fusión en individuos. Abundan los detalles que apoyan el concepto de que todos los organismos visibles —plantas, animales y hongos por igual— evolucionaron por medio de la «fusión de cuerpos». La fusión a nivel microscópico condujo a la integración genética y a la formación de individuos cada vez más complejos. El impulso termodinámico hacia la aparición de sistemas de reducción de gradientes cada vez más complejos encuentra su expresión en la creación continua de nuevas y cada vez más intrincadas formas de asociaciones entre formas de vida, entre las que figura la simbiosis.
§. La fuerza del número
Terry Erwin, profesor de la Universidad de Alabama que hace caer insectos de la bóveda amazónica para contarlos, nos asegura que, como norma general, dos tercios de las especies con las que se encuentra son nuevas para la ciencia. Erwin sugiere que, si hasta la fecha se cuentan diez millones de especies documentadas en los anales académicos, y si los números que él encuentra representan a la totalidad del planeta, deberían existir sobre la Tierra al menos treinta millones de especies de animales vivos. Joseph Leidy, un sabio de la Filadelfia del siglo XIX y uno de los fundadores de la Academia de Ciencias Naturales de esa ciudad, resumía así su filosofía de una Buena Vida: ¿Cómo podría la vida ser aburrida, quedando todavía un rizópodo más por describir? Tanto si la medimos por los rizópodos (amebas) de Leidy, como por los escarabajos de Erwin o los millares de especies de orquídeas de las montañas de Colombia, la diversidad de la vida deja pasmada a la imaginación. La tarea linneana de clasificar todo lo vivo está tan inacabada en el siglo XXI como lo estuviera en el XVII, cuando la señora Linneo tuvo que vender toda la colección de su marido a Londres para poder pagar a sus acreedores y financiar la dote de su hija. Los especímenes de sus tarros, sus botellas y sus plantas secas siguen reposando plácidamente en el sótano de la Sociedad Linneana en Burlington House, muy cerca del bullicioso Piccadilly Circus.
El objetivo de catalogar las especies vivas sobre el planeta sigue siendo igualmente noble, de hecho más noble y útil si cabe que en tiempos de Linneo, cuando las especies eran catalogadas y clasificadas de forma práctica, sin tenerse que preocupar por la conservación de la biodiversidad del planeta. Linneo nunca creyó que las formas de vida fueran el reflejo de una historia evolutiva, sino que pensaba que toda aquella abundancia y diversidad biológica eran obra de una deidad buena y prodigiosa. Su concepto de especie pertenecía a una categoría fija e inmutable, identificable por sus características aparentes. La mayor parte de los nombres que puso a sus 10.000 especies siguen en uso hoy en día.Phantera leo es el nombre que le dio al león yFelis catus el que le puso al gato común. A las manzanas las llamó Malus deliciosus y a los exquisitos mejillones de la costa atlántica les puso por nombre Mytilus edulis. El primer término, siempre en mayúscula, se refiere al grupo mayor o «clase» de forma viva (personas, perros, manzanas, mejillones, etc.), es decir al género, mientras que el segundo término o «epíteto específico» indica la especie (personas sabias, perros cariñosos, manzanas deliciosas, mejillones comestibles, etc.). Y lo mismo en lo que se refiere a los hongos —Lactarius deliciosus (exquisita seta lechosa) o Penicillium chrysogenum (pequeño lápiz dorado)— que, puesto que no eran animales, según Linneo tenían obviamente que ser plantas.
En resumen, las especies eran y siguen siendo el mínimo denominador común. Desde los tiempos de Linneo hasta nuestros días, las especies han ido engrosando el «banco» de la bibliografía: el nombre, el lugar de hallazgo, la descripción publicada y los datos de su autor se depositan públicamente en un herbario o en un museo de historia natural. A medida que se generalizaba el uso del microscopio, proliferaban las especies y las prácticas de dar nombre se extendían para incluir a las bacterias (como plantas, aunque nadaran) y a otros organismos unicelulares como plantas si eran verdes o verduscas, o como animales si nadaban.
Sobre la definición de las especies se han escrito muchos millones de palabras. Aquí proponemos una idea verificable acerca de lo que las especies son.
§. Las especies desconstruidas: la ardisia de miehe y la euplotidium de rosati
La existencia y extinción de especies, así como el origen de otras nuevas, constituyen fenómenos que encontramos únicamente en los organismos nucleados. Este nuevo concepto no es más que un simple corolario a la idea de Sonea de que las poblaciones procariotas no forman especies en la Naturaleza. Los procariotas, por el contrario, tanto si se trata de arqueobacterias como de eubacterias, pertenecen a una única especie global. ¡El Internet bacteriano precedió al nuestro en unos 3.000 millones de años [11]! Los organismos con organización celular bacteriana no son clasificables en especies. Los organismos nucleados, producto todos ellos de la simbiogénesis, aparecieron por primera vez sobre la Tierra hace más de 1.200 millones de años. Su aparición se corresponde enteramente con la de las primeras especies y los primeros géneros. ¿Por qué? En nuestra opinión porque: Un organismo (A) pertenece a la misma especie que otro organismo (B) cuando —y sólo cuando— A y B tienen precisamente los mismos antepasados celulares, es decir, descienden de los mismos genomas y las relaciones entre esos genomas son idénticas. En otras palabras, si hacemos una lista con los genomas que constituyen A y otra con los que constituyen B, ambas listas serán idénticas cuando ambos organismos pertenezcan a una misma especie, y diferirán si pertenecen a especies distintas.
Desde hace tiempo, los zoólogos han reconocido la validez del concepto biológico de especie de Ernst Mayr: dos organismos pertenecen a la misma especie cuando se reconocen mutuamente, pueden aparearse y pueden producir descendencia fértil. Estamos de acuerdo con esta definición, que resulta apta para la mayoría de los animales y para muchas plantas. En realidad, la vemos como un caso particular de nuestra definición. Únicamente los animales que comparten un mismo complemento de genomas pueden desarrollar géneros complementarios que puedan permitirse apareamientos fértiles. Sin embargo, nuestra definición reconoce la composición heterogenómica de todos los organismos nucleados, por lo que resulta ser mucho más amplia. Nuestro planteamiento de «lista de componentes de especies» admite la existencia de decenas de millares de especies de protoctistas, de hongos, de plantas y de animales que no se aparean para producir descendencia.
A lo largo de este libro revisaremos algunos ejemplos de nuevas especies y nuevos taxones que se originaron por medio de la adquisición y la incorporación de genomas previamente independientes. Veamos a continuación un par de ejemplos sobre cómo analizar las especies a partir de sus componentes genómicos.
§. El secreto del borde de la hoja de miehe confirmado
Las especies y los géneros nuevos de plantas de flor evolucionaron cuando las hojas de estas plantas adquirieron e integraron a su historia vital un genoma bacteriano. En la familia de las plantas de flor denominada mirsináceas se encuentran tres géneros localizados en Indochina, Formosa, Malaca, la península de Malaya y las Filipinas. Los miembros del género Ardisia, el más extendido de los tres, tienen hojas alabeadas (figura 3.1).
Figura 3.1 Borde de hoja de Ardisia, alabeado por la acción de colonias de bacterias.
La mejor conocida de entre las treinta especies de Ardisia es la A crispa. Todas ellas, junto con cinco especies de los géneros menos conocidos, exhiben en sus hojas simbiontes bacterianos regulares. DeJong sugirió que esta asociación se habría originado a partir de las bacterias próximas del suelo, tal vez en una única ocasión (monofiléticamente). Según DeJong, la simbiosis habría arraigado porque las plantas crearon dependencia de las substancias promotoras del crecimiento generadas por las bacterias. Las bacterias que forman las hojas alabeadas de la Ardisia crispa están tan perfectamente integradas en la planta, que se las toma simplemente como la característica que distingue a las hojas de esta especie.
Sugerimos que esta simbiosis bacteriana integrada se correlaciona tanto con la morfología de las hojas como con el establecimiento de nuevas especies y nuevos géneros de Ardisia. La diferenciación de especies entreYa Ardisia crispa y los demás miembros distintos del subgénero Crispardisia está directamente relacionada con la adquisición e integración de simbiontes bacterianos parecidos pero no idénticos.
§. Ciliados defensivos o la revelación de rosati
Veamos otro ejemplo. En las aguas costeras del Adriático medran los ciliados, protoctistas monocelulares que forman claramente parte de la gran familia de ciliados denominada «heterotricoides». “Hetero” significa otro y “tricoides” parecido al pelo. El nombre se debe a las protuberancias móviles de sus cuerpos (figura 3.2.) al igual que otros ciliados heterotricoides, éstos tienen «bocas» bordeadas por filas de cilios denominados membránulas, que barren hacia la boca su alimento bacteriano. El ciliado nadador es asimétrico, su dorso o espalda está cubierto por menos grupos de cilios (denominados cirros) que su parte ventral o frontal.
El género de los Euplotidium consta de seis especies, todas las cuales se cree que evolucionaron a partir de un antepasado común, denominado Euplotes, género cosmopolita de ciliados heterotricoides que contiene centenares de especies, todas ellas bacterívoras, es decir, que se alimentan de bacterias. Los Euplotes son mucho mejor conocidos que los Euplotidium, aunque ambos sean parecidos en cuanto a aspecto externo se refiere.
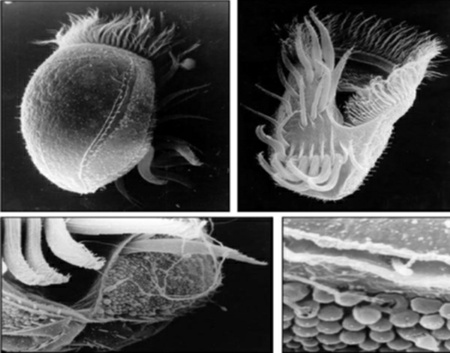
Figura 3.2 Las «granjas corporales» de los Euplotidium, sus órganos defensivos.
La transformación desde la fase primera a la segunda está plenamente correlacionada con el ciclo celular del ciliado sobre cuya superficie están embebidas estas bacterias. Cuando el ciliado detecta en las aguas del Adriático indicios de peligro en forma de presencia de predadores potenciales, tales como otro ciliado, el Litonotus lamella, se produce una violenta reacción: el disparo de las cintas enrolladas de la banda de bacterias de la superficie del Euplotidium. Centenares de bacterias proyectan al unísono sus peligrosas cintas hacia fuera, alcanzando una longitud de cuarenta micrones, casi tan grande como la del propio ciliado, a pesar de que la estructura en la que la cinta estaba enrollada no superara un micrón. Los dos organismos simbióticos actúan de este modo como uno solo: cuando el ciliado se siente amenazado por predadores o por otras perturbaciones, un número masivo de las bacterias que viven en su superficie proyectan sus cintas.
La asociación entre el ciliado Euplotidium, en todas sus seis especies, y las bacterias disparadoras de cintas es constante. Todos los miembros del género exhiben esta misma ectosimbiosis protectora. En el laboratorio es posible retirarles esta protección. Sin sus bandas de bacterias protectoras, los ciliados nadan, se alimentan y se reproducen por división como de costumbre. Pero su predador Litonotus lamella permanece siempre al acecho en busca de un Euplotidium que llevarse a la boca. Una vez desprovisto de su armadura de bacterias ectosimbióticas lanzadoras de cintas, el Euplotidium se convierte en presa fácil para el Litonotus lamella, que no tarda en devorarlo. Cabría argumentar que la presencia de esas bacterias protectoras sobre la superficie del Euplotidium impide la selección natural. Sugerimos que la adquisición de estas bacterias y su integración como orgánulos por algún antepasado parecido al Euplotes condujo a la aparición de una nueva especie, el Euplotidium itoi. Puesto que otras cinco especies de Euplotidium incluyen bacterias protectoras, parece probable que el género Euplotidium se originara en los Euplotes a partir de la adquisición y la integración de genoma microbiano. Hoy en día, las bacterias lanzadoras de cintas del E. itoi son orgánulos extracelulares, sin los que el ciliado no puede vivir en el medio natural. Tampoco esas bacterias pueden cultivarse independientemente, sino únicamente en asociación con el E. itoi, es decir, que su existencia misma depende del ciliado al que están vinculadas. Los miembros del género Euplotidium existen, pues, únicamente como entidades simbióticas; ninguno de los dos organismos que las componen es viable por separado. Nos encontramos, por consiguiente, ante un ejemplo claro, presumiblemente bastante reciente, de origen de una especie debido a la herencia de genomas adquiridos.
§. Comunidades y la lógica de los ecosistemas
Los miembros de una misma especie que viven juntos, al mismo tiempo y en un mismo espacio, constituyen poblaciones. Todos los organismos viven en comunidades formadas por poblaciones. Todas las comunidades están formadas por dos o más especies distintas que conviven en hábitats identificables. Esta organización se relaciona con precisión en la Naturaleza con factores ambientales tales como el clima y la geografía, entre otros. La taxonomía —la ciencia que se ocupa de nombrar, identificar y agrupar a los seres vivos— tiende a ignorar las correlaciones medioambientales y es por tanto artificial, particularmente en lo relativo a las bacterias. Aun así, en los casos de los protoctistas, los hongos, las plantas y los animales, los grupos de nivel más fácilmente identificados y nombrados son los del género y la especie. A pesar de la advertencia zoológica de que las especies deben establecerse en función de la capacidad de sus miembros para aparearse y producir descendencia fértil, las especies se distinguen, se cuentan, se documentan y se nombran principalmente por cuestiones de apariencia y morfología.
Aunque sean reconocibles diversos tipos de bacterias, éstas son mucho menos estables que los organismos superiores en cuanto a características se refiere. Las «especies» bacterianas son tan evasivas e inidentificables que, en realidad, el concepto de especie no se les puede aplicar. Hay quienes llaman «ramas» a los grupos bacterianos, dejando la discusión de los detalles para mejor ocasión. Cualquier bacteria puede pasarle genes a otra. Las restricciones en este flujo promiscuo de genes, y por consiguiente la posibilidad de especiación, comenzaron en el Eón Proterozoico inferior, hace unos 2.500 millones de años, cuando las transformaciones de células bacterianas en consorcios y comunidades condujeron a la integración, a la construcción de contornos y, finalmente, a las primeras células eucariotas. Estas células, como hemos explicado en otro lugar, son en sí mismas disposiciones simbióticas. Partes de las células eucariotas tales como las mitocondrias, e incluso tal vez los cilios y sus microtúbulos, evolucionaron originariamente a partir de organismos independientes. Así pues, la simbiogénesis explica el origen de las especies en dos sentidos: en primer lugar reuniendo las bacterias en el pasado pretérito para dar lugar a organismos susceptibles de especiación; en segundo lugar, y como veremos, creando nuevas especies por medio de la incorporación de otros cuerpos y, finalmente, de genomas. Contenido:
La reproducción o la voluntad de poder de Nietzsche§. La reproducción o la voluntad de poder de Nietzsche
Poblaciones incontroladas
El reconocimiento de Darwin
Gaia como selector natural
Todos los organismos llevan dentro de sí el equipo químico necesario para la reproducción. Las bacterias se dividen en dos, dos se convierten en cuatro, cuatro en dieciséis y así ad infinitum. Los foraminíferos, grandes protoctistas con cáscara que viven en el océano, se multiplican por centenares. Un solo progenitor puede engendrar 320 pequeños descendientes nadadores en cuestión de pocos minutos. En los animales y las plantas, el apareamiento es a menudo el único juego disponible. Con o sin pareja, la reproducción constituye el impulso final, el objetivo de la vida. Friederich Nietzsche describió la tendencia incesante de la vida a seguir su propio curso, a crear y expandirse, a ver las cosas según cierta interpretación poética que tiende a ser olvidada, como «la voluntad de poder».
En contraposición a esta fuerza creativa está la selección natural, que no genera nuevas formas, ni innova ni produce, sino que —como su nombre indica— simplemente selecciona entre lo previamente creado.
¿Para qué sirve pues la elección de la selección natural? Del mismo modo que muchos evolucionistas modernos se permiten cierta vaguedad poco científica acerca del papel de la selección natural en la evolución, también se otorgan licencia para mostrarse igualmente ambiguos acerca de la identidad del selector natural. Es demasiado fácil levantar los hombros y afirmar que «el entorno selecciona; los más aptos sobreviven». Para empezar, ¿qué significa exactamente “apto”? ¿Qué partes del entorno son las que seleccionan? ¿Hasta dónde llega el entorno? Las preguntas de esta índole tienden a ser contestadas ambiguamente o por medio de respuestas puntuales, caso por caso. En seguida asoma una férrea resistencia a cualquier esfuerzo sistemático de identificación del agente o los agentes de la selección natural.
Declaramos en este capítulo la sencilla pero importante afirmación de que el selector natural es Gaia, entendiendo por tal a la biosfera, es decir, todo el conjunto de interacciones que se dan sobre la superficie de la Tierra junto con la biota o materia viva, organizadas de forma natural en ecosistemas. La «biota», o biomasa total, se refiere a la flora, la fauna y la microbiota como un todo. La selección natural constituye uno de los medios por los que Gaia, como sistema autorregulador, se mantiene a sí misma como una entidad dinámica y al mismo tiempo estable.
§. Poblaciones incontroladas
Los constreñimientos medioambientales cotidianos, como la falta de agua, la masificación o el hambre, previenen la expansión indefinida que las poblaciones seguirían de por sí. Puesto que cada población tiene sus energías específicas —carbono, nitrógeno, agua, etc.—, su espacio y otros requerimientos, que nunca son completamente proveídos por el entorno, la expansión de la población se ve inevitablemente limitada. La selección natural —proceso netamente sustractivo— elimina a quienes, por una u otra razón, no logran sobrevivir para reproducirse. Por definición, los que quedan sobreviven y tienden a transmitir a su progenie sus rasgos heredables. Habida cuenta de que estos descendientes retendrán rasgos conducentes a la supervivencia en determinados tiempos y lugares, la vida sobre el planeta guarda en cierto modo el registro de su pasado. En sus complejas memorias químicas, los cuerpos vivos almacenan registros de las limitaciones ambientales del pasado que lograron superar.
§. El reconocimiento de Darwin
¿Por qué alberga una sola termita treinta especies distintas de protistas y más de 200 clases distinguibles de bacterias? ¿Por qué en el ecuador de los trópicos, donde el Sol sale exactamente a las seis de la mañana y se pone exactamente a las seis de la tarde, donde las condiciones de vida son tan tremendamente uniformes, son los seres vivos tan espectacularmente diferentes? ¿Por qué en los picos pelados de los volcanes de las islas Galápagos, desoladas bajo un sol incesante y tan parecidos en tamaño, historia, agresiones físicas y geografía, son sus habitantes tan profundamente diversos? «Esto se me ha antojado siempre como una gran dificultad —escribió Darwin hacia el final de suOrigen de las especies según cita Jonathan Wiener en The Beak of the Finch— que procede en gran medida del error profundamente arraigado que consiste en considerar a las condiciones físicas del entorno como lo más importante para sus habitantes, cuando creo que ya no se puede discutir que la naturaleza de los otros moradores, con los que cada cual debe competir, constituye un elemento al menos tan importante, si no más, para el éxito».
Que un nuevo ser vivo empollado, parido o germinado prospere, viva, se multiplique y sobreviva dependerá en parte, por supuesto, de las condiciones climáticas y geográficas de su entorno, pero que acabe convirtiéndose en alimento para otro o en su amigo y aliado en una gran batalla, o que reciba sombra o calor o sufra por el contrario de la ira de cualquier otro, dependerá en gran medida de la benignidad o de la agresividad de esos otros seres vivos, con los que compartirá ese entorno en el que le ha tocado tratar de sobrevivir. Sea cual fuere su naturaleza, que los seres vivos lleguen o no a reproducirse depende de los otros seres que vivan en su entorno inmediato, sean éstos de otra especie o de la suya propia.
§. Gaia como selector natural
Gaia, el nombre de la antigua diosa griega de la Tierra, utilizado ahora como nombre de una teoría para el planeta vivo, se define tal como describe la leyenda de la figura i.
El término “biota” se refiere a la totalidad de la flora (plantas), la fauna (animales) y la microbiota (hongos, protoctistas y bacterias), es decir, equivale a la suma total de toda la biomasa del planeta; en otras palabras: a toda la materia viva presente. La biota comienza y acaba dentro de la biosfera, es decir, de la parte de la esfera terráquea donde mora la vida y que abarca desde la profundidad de las simas oceánicas hasta la troposfera. En ella viven más de treinta millones de tipos de organismos, especies y cepas bacterianas, descendientes todos ellos de antepasados comunes. Todos interactúan entre sí. Todos producen y eliminan gases, iones, metales y compuestos orgánicos. El metabolismo, el crecimiento y las interacciones de esta miríada de seres, particularmente dentro de soluciones acuosas, condujo a la modulación de la temperatura, la alcalinidad y la composición atmosférica de la superficie terrestre.
Está, pues, claro que Gaia se refiere a la suma de todos los seres vivos, más que a un organismo “A” arbitrariamente elegido, inmerso en su propio entorno sobre la superficie del globo. La visión gaiana de la vida incluye también el entorno de todas esas otras formas de vida. Es Gaia, como un todo, la selectora de ese organismo “A” como miembro de la población de “A” dentro de la Naturaleza. Es Gaia, en general, quien previene que las poblaciones del organismo “A” desarrollen todo su potencial. Como en toda población, desde las bacterias del ántrax en un pulmón caliente a las ratas de Nueva York en los sótanos de las casas viejas, el crecimiento por medio de reproducción seguirá y seguirá hasta ser detenido… por la selección natural. El hecho de que Gaia seleccione de forma natural nos ayuda a comprender por qué la Tierra es un sistema integrado.
Capítulo 5
Principios de novedad evolutiva
Fuentes de novedad heredableEl desarrollo de superpoderes a través de mutación debida a la química o a la radiación, dando pie teóricamente a la aparición de nuevas especies, constituye un fenómeno fácil de encontrar en muchos cómics. Las mutaciones se definen como diferencias entre progenitores y descendientes, susceptibles de ser transmitidas genéticamente a las generaciones posteriores. Pueden ser espontáneas o inducidas por tratamiento. Al exponer a radiaciones de rayos X a moscas de la fruta, H. J. Muller contribuyó al descubrimiento de las bases químicas de la mutación. Muchos otros científicos han aplicado con éxito agentes mutagénicos, ya sea por radiación o con ayuda de la química, a gran cantidad de organismos. Si bien se han registrado cambios heredables, nunca se ha logrado dar pie a una nueva especie. Los fármacos para el tratamiento del cáncer son mutagénicos, pero en ningún caso han conducido a la aparición de una nueva especie. Algunos cambios en los pares base del ADN, así como algunas mutaciones cromosómicas, también han producido pequeños cambios heredables.
En cambio, la mutación por sí misma suele generar organismos enfermos y formas de vida deficientes. Los cambios mutacionales favorables son siempre pequeños. Las nuevas mutaciones generan variaciones en miembros de una misma especie pero, ni en el laboratorio ni sobre el terreno, la acumulación de mutaciones ha logrado nunca saltar las barreras entre especies. Han tenido lugar muchos intentos de demostrar los efectos de la mutación sobre la evolución. Tal vez el más reciente sea el de Barbara Wright con bacterias. Dadas unas condiciones altamente específicas, las mutaciones no son aleatorias y, en determinados entornos, son recogidas por las bacterias descendientes (Wright, 2000).
A diferencia de la mutación, la rápida adquisición de rasgos nuevos y refinados por medio de la adquisición e integración de organismos precedentemente extraños confiere ventajas inmediatas a sus captores protoctistas, plantas o animales por igual. A menudo la asociación comienza de forma predatoria: uno de los organismos trata de ingerir y digerir al otro, que se resiste, la presa atrapada o la bacteria no digerida dan origen a una población cautiva. La herencia de poblaciones cautivas, particularmente en forma de genomas microbianos, crea nuevos linajes evolutivos que exhiben estrategias genuinamente novedosas merecedoras de estudio. Sin duda ocurren mutaciones aleatorias —tanto en el predador como en la presa— y por supuesto estas mutaciones constituyen aspectos importantes del proceso evolutivo. Pero las mutaciones refinan y aguzan. En los eucariotas, las mutaciones por sí solas no crean nuevas especies ni variaciones heredadas positivas de importancia. En cambio, los genomas vienen limpiamente empaquetados con dilatadas historias de virtuosismos y trucos sintéticos heredables. Proporcionan precisamente lo que hace falta para que un organismo pueda cambiar drásticamente sin dejar por ello de seguir siendo coherente y viable. Documentación fiable de la literatura científica demuestra que la movilidad, la capacidad para transformar alimentos o la aparición de nuevos rasgos metabólicos (producción de vitaminas, extracción de nitrógeno a partir del aire, destoxificación del oxígeno) pueden ser apropiados como un todo, como en el caso de la absorción de los empleados expertos de una empresa absorbida por otra.
Justamente del mismo modo que tras un casamiento o una fusión empresarial no es sencillo dar marcha atrás, la evolución por medio de adquisición genómica constituye un proceso irreversible. Tras la integración de genomas más pequeños en los genomas del organismo receptor, el genoma anteriormente externo ya no puede ser liberado de nuevo a la Naturaleza. Llega un punto en la relación en que la forma de vida que antes vivía libre ya no puede ser voluntariamente rechazada o «desadquirida». Existen ejemplos de rechazo de socios de largo recorrido, pero todos ellos tienen como resultado la aparición de nuevos seres modernizados y alterados, nunca la reversión a las formas pasadas.
Lamarck se equivocaba al afirmar que los cambios de comportamiento en la generación progenitora conducían a cambios heredados en sus descendientes. En cambio, a través de la adopción y subsiguiente integración de genomas, en una sola vida pueden ser adquiridos nuevos y espectaculares rasgos. En nuestro libro anterior (Microcosmos, 1997) describíamos el único experimento de laboratorio con resultado de aparición de una nueva «especie»: la infección permanente de amebas estudiada por Kwang Jeon de la Universidad de Tennessee. En determinados casos, los cambios en el comportamiento y en el cuerpo pueden ser heredados cuando son causados por la herencia de un genoma bacteriano o vírico. He aquí un claro ejemplo: las hojas de la Abutilón pictum thompsonii, una variedad de planta cuyas manchas son causadas por un virus y que mostramos en la figura 5.1. En tales casos Lamarck tal vez no estuviera demasiado errado. El rasgo en sí mismo no es heredado, pero los genomas adquiridos que lo determinan sí lo son. Frotar o rascarse los genitales promueve la transmisión de microbios íntimos, como la espiroqueta de la sífilis o el protista ( Trichomonas vaginalis) que produce picor vaginal. Estornudar disemina organismos y virus respiratorios. Las lombrices verdes que albergan algas simbiontes buscan la luz, cosa que no hacían sus antepasados blancos. El efecto de la adquisición de simbiontes sobre el comportamiento es una cuestión digna de investigación por los evolucionistas serios.

Figura 5.1 Jaspeado vírico: Abutilón pictum (Malváceas).
¿Cómo consigue un organismo adquirir nuevos genes completos? Las bacterias predadoras se tragan a sus presas. Otras bacterias activamente comedoras producen babaza, polisacáridos blanquecinos que actúan como adhesivo. Otras son engullidas por protistas. Estos comportamientos conducen a la adquisición, a la unión y en ocasiones a la fusión. Las simbiosis más recientes, como la de la bacteria que se alimenta entre las células ( Aeromonas hydrophila) o los ciliados que recorren las superficies húmedas en busca de bacterias y otros nutrientes en la Hydra viridis, nos ayudan a comprender cómo se iniciaron asociaciones más profundas y perdurables. El Streblomastix strix, un protista que navega por el intestino de las termitas, está recubierto por pelos sensitivos que reconocen el acetato que le sirve de alimento. Estos pelos resultan ser bacterias colgantes. Al serle retirados por medio de antibióticos, el Streblomastix ya no se sentía atraído por el acetato. En el sentido evolucionista, los protistas tendrían menos posibilidades de sobrevivir sin sus órganos sensoriales formados por bacterias heredadas.
Los plástidos verdes envueltos en membranas de las algas fueron otrora bacterias duras y resistentes que les sirvieron de alimento. Las termitas caníbales retienen en su propio intestino a los simbiontes de sus compañeras ingeridas. Los nematocistos —dardos venenosos naturales construidos a partir de partes de células— de las babosas marinas están tomados de los celentéreos que se comen, para ser usados como medio de defensa de la propia babosa. Todos estos ejemplos no tan sólo nos demuestran cómo los simbiontes integrados tienden a seguir adquiriendo nuevos simbiontes, sino que también ponen de relieve que la historia simbiótica es destino evolutivo.
Habida cuenta de que la supervivencia diferencial es todo lo que hay, la selección natural perpetúa pero no crea. ¿Qué es entonces lo que genera innovación evolutiva? La relación de fuentes de cambio hereditario —mutación, en el sentido más amplio del término— se hace cada vez más extensa, dentro de la cual la adquisición e integración de genomas es un apartado más. La novedad aparece y se acumula por un simple cambio en un par de bases del ADN, sea éste aleatorio o no. Las mutaciones del ADN pueden inferirse fácilmente a partir del estudio directo de las secuencias de aminoácidos en las proteínas, ello debido a que dicha secuencia viene codificada por la de las bases en los genes. No hay que esperar copias perfectas en un cosmos regido por la segunda ley de la termodinámica. Los organismos adquieren también nuevos rasgos hereditarios por acumulación de virus, de plásmidos o de otros fragmentos cortos de ADN. También pueden hacerse con grandes fragmentos de ADN nuevo —muchos genes a la vez— mediante apareamiento bacteriano o por relación sexual legítima con parientes lejanos, es decir, por hibridación. Las modalidades de construcción de variedad por adquisición no genómica, es decir, por intercambio de genes uno por uno, son desproporcionadamente importantes en el universo de las bacterias. Tomadas como un todo, estas modalidades constituyen un serio argumento en la resolución del dilema de Darwin. La ciencia sí sabe hoy cuáles son las principales fuentes de novedad evolutiva. La tarea consiste ahora en sacar a este conocimiento de los tomos de la esotérica y darlo a conocer mejor.
¿Cómo se adquiere el nuevo material genético? ¿Qué cantidad de material genético nuevo es susceptible de ser adquirido gen por gen, o unos cuantos genes cada vez, únicamente aquellos que se desee adquirir? La nueva tecnología de secuenciación del ADN nos permite responder a estas preguntas de forma directa.
El mínimo cambio genético heredable consiste en el cambio de uno de los pares de bases del ADN, de A-T a G-C o a la inversa. A su vez, el máximo cambio genético heredable posible consiste en la adquisición de la serie completa de genes que regulan el funcionamiento de un organismo —su genoma—, junto con el resto de ese organismo en buen estado de salud, de forma que el genoma adquirido siga teniendo un funcionamiento que regular. Entre ambos extremos existe un sinnúmero de modos en que los organismos consiguen y retienen novedad heredable. Cuando, a principios del presente milenio, fue anunciada la secuencia completa del genoma humano, fueron numerosos quienes se sorprendieron de saber que cerca de 250 de los más de 30.000 genes humanos procedían directamente de bacterias. El origen bacteriano de estos genes, largas secuencias de ADN que codifican las proteínas, es tan fácilmente reconocible como lo pueda ser el de una pluma de ave frente a, por ejemplo, la boca de un tiburón.
El genoma de la levadura Saccharomyces cerevisiae ha sido completamente secuenciado, con el resultado de una buena sorpresa para los científicos. Eligieron secuenciar esta levadura como representante del reino de los hongos, porque la vida de su versátil y pequeña célula está relacionada con la nuestra de múltiples formas. Esta levadura es la que monta la masa y, por consiguiente, la mayoría de alimentos cocidos al horno —pan, pasteles, brioches…— dependen de ella. Es asimismo la responsable de la fermentación la cerveza, por lo que cualquier cerveza con alcohol depende de su feliz desarrollo. Abunda igualmente en el yogur y en otros productos lácteos. En el laboratorio crece bien y rápidamente, habiendo sido por ello, desde hace tiempo, la favorita para el estudio del sexo de los hongos, de los virus fúngicos, del comportamiento, crecimiento y supervivencia de los cromosomas, y de la formación de esporas. Como es bien conocido, la célula de la Saccharomyces cerevisiae es «haploide», es decir, tiene una sola copia de cada uno de sus diez cromosomas (los humanos somos diploides, lo cual significa que cada uno de nosotros tiene dos copias de cada uno de los veintitrés cromosomas: un juego procede de la madre y el otro del padre). La sorpresa a la que nos referíamos consistió en el descubrimiento de que la levadura tiene dos copias de casi todos sus genes.
La historia de la levadura añade otro capítulo a nuestra colección de formas de adquirir material genético nuevo: la duplicación. Cada uno de los organismos que han sido estudiados presenta algún grado detectable de duplicación de genes: una parte de un gen antiguo, un solo gen completo, un conjunto de genes nuevos, un cromosoma entero, o —como en el caso de la levadura— casi todos los genes contenidos en el cuerpo de la pequeña célula. Del mismo modo que las copias extra de manuscritos originales o de libros de instrucciones aseguran que el original no difiera de sus copias, los conjuntos de genes extra han demostrado ser muy útiles cuando las levaduras u otros organismos han evolucionado hacia mayor tamaño o creciente complejidad.
Parte II
Los microbios en la evolución
Reflexionando sobre el consenso entre los evolucionistas de nuestros días, Wiener trata de resumir un problema que el propio Darwin enfrentó con confusión y ambivalencia: la relación entre la selección sexual y la selección natural. Wiener propone como ejemplos a dos prominencias humanas: la primera, la fatalmente atractiva nariz de Cleopatra (seguida, por supuesto, del resto de su rostro y de su cuerpo), que condujo a hombres como Julio Cesar y Marco Antonio a acciones que determinarían cambios en la estructura de los pueblos del Mediterráneo. La segunda, la nariz del propio Darwin, descrita por
Robert FitzRoy, primer oficial ejecutivo del Beagle, como la de un hombre perezoso. Si el Capitán FitzRoy se hubiera abstenido de enrolar aquel joven naturalista en la aventura del Beagle hacia las costas sudamericanas, el resto de los mortales no estaríamos hoy debatiendo sobre El origen de las especies. Tal vez en lugar de ello estaríamos ponderando el trabajo seminal de Alfred Russel Wallace o de T. H. Huxley. Incluso el impulso erótico de algún marinero británico, inspirado por los senos colgantes y el escaso y tieso vello púbico de las nativas de las islas Salomón, puede tener repercusiones evolutivas, si esa atracción tiene como resultado la aparición de una nueva subespecie de nativos de las islas del Pacífico. Como también las puede tener un huracán brutal, si como resultado no quedan vivos más que dos adolescentes de piel oscura y nariz achatada para repoblar las cimas de esos conos volcánicos que los nativos polinesios llaman su hogar. La selección sexual y la selección natural son variaciones de un mismo tema: tan sólo una ínfima porción de entre una gran diversidad sobrevive para propagar su estirpe.
El obstáculo, la dificultad, la causa de desasosiego y de exceso de verborrea, no es otra que la afirmación —o suposición tácita, o «dato» de libro de texto— de Darwin y sus seguidores de que esta clase de selección conduce a la aparición de nuevas especies. La pesada mano de la selección puede ejercer y ejerce cambios tremendos en la proporción de huevos fértiles, de gallinas ponedoras, de tomates grandes o pequeños, o de pinzones terrestres de pico largo. Pero nadie ha podido nunca demostrar que este proceso haga algo más que cambiar las frecuencias genéticas en las poblaciones. La variación intraespecífica no parece haber desembocado nunca, por sí sola, en nuevas especies.
Como estudiante en los años 60, Niels Eldredge, que con el tiempo se convertiría en el responsable de zoología invertebrada del Museo de Historia Natural de la ciudad de Nueva York, se dedicó a buscar en los registros fósiles pruebas directas de especiación. Estudió con ahínco colecciones enteras de trilobites cámbricos primorosamente conservados, en busca de transiciones graduales de una especie a sus especies descendientes. Tanto en Marruecos como en el estado de Nueva York, peinó cuidadosamente los sedimentos en secuencias estratigráficas. Halló, de capa en capa, algunas variaciones en el tamaño y en la forma del caparazón, pero en ningún caso encontró alguna tendencia clara que indicara una lenta transición entre una especie y otra. Más bien parecía que la presencia de la misma especie proseguía, con pequeñas variaciones aleatorias, a lo largo de 800.000 años. De repente aparecía otra, que superaba a la anterior en 1,3 millones de años. La búsqueda de formas intermedias y de cambio evolutivo gradual entre ambas demostró ser siempre fútil. Las rocas sedimentarias en las que duermen los gloriosos registros fósiles no mienten ni engañan. El registro era puntuado, las diferencias entre especies de animales extintos atrapadas en el tiempo eran claras y perfectamente distinguibles. Las pequeñas variaciones dentro de una misma especie, indicativas de cambios en la frecuencia de sus genes, oscilaban arriba y abajo sin dirección aparente («equilibrio» dentro del «equilibrio puntuado»). La aparición de especies y géneros nuevos, así como la pérdida de otros por extinción demostraban ser siempre discontinuas (ahí reside la «puntuación»).
Las conclusiones de Eldredge con sus trilobites, confirmadas más tarde por Richard Fortey y sus colegas, se repitieron con caracoles de las Bahamas en su trabajo sobre la «teoría del equilibrio puntuado», realizado junto con Stephen Jay Gould. El mismo registro fósil ha aparecido en yacimientos de moluscos y de dinosaurios, así como en la historia reconstruida a partir de los restos óseos de animales marinos gigantescos que aterrorizaron a otros pobladores de los mares del Devónico. Cualquiera que haya sido testigo de las dificultades de lograr descendencia entre una hembra dachshund y un pastor alemán macho, sabrá lo que son las variaciones intraespecíficas. Sin embargo, nadie que haya estudiado cuidadosamente los organismos de ambos animales en su estado natural habrá podido detectar ningún cambio gradual de una especie a otra, ni en sus poblaciones vivas ni en sus restos fósiles. Cuando las investigaciones se aproximan a los límites entre especies, los abismos genuinos —entre rábanos y coles, entre asnos y caballos, o entre bonobos y chimpancés— siguen ahí. Al menos entre animales fácilmente reconocibles, los límites están claros.
§. Las bacterias como especie única
Las bacterias no forman especies. Sus diferentes tipos cambian constantemente con fluidez y rapidez. Comparado con las plantas o con los animales, el repertorio metabólico de las bacterias es inmenso. En sus descendientes protoctistas, el metabolismo es más limitado, pero aún así sigue siendo más extenso que el de plantas, animales y hongos. Mucho antes de la aparición de formas de vida fúngicas, animales o vegetales, el universo bacteriano había ya refinado exquisitamente la elección, el desarrollo, la modalidad de apareamiento, la inmunidad, la formación de minerales, el movimiento, la fotosíntesis, la predación, la muerte celular programada, la sexualidad y un montón de otros atributos de vida «superiores». Tan sólo recientemente hemos comenzado a apreciar las verdaderas dimensiones del pasmoso repertorio metabólico, así como de la enorme diversidad morfológica del reino de las bacterias. (En nuestro caso, hemos llegado incluso a argumentar que, tanto en el mundo de las bacterias como en el de sus descendientes protoctistas, se habría originado algo parecido a la libre elección, lo cual nos valió que el biólogo celular y premio Nobel Christian Duve nos acusara en las páginas de la revista Nature de «biomisticismo»).
Una prodigiosa bibliografía técnica demuestra que las bacterias constituyen la mayor reserva de diversidad evolutiva. Todo el reino animal emplea esencialmente una misma modalidad de metabolismo: la utilización del oxígeno para respirar moléculas orgánicas de alimento. Esta modalidad se conoce con el nombre de heterotrofia. Las plantas utilizan dos modalidades: la heterotrofia vía oxígeno —como los animales— y la fijación de dióxido de carbono y la fotosíntesis productora de oxígeno, utilizando para ello la radiación solar, que se conoce con el nombre de fotoautotrofia. Además de estas dos modalidades —heterotrofia y fotoautotrofia—, las bacterias disponen como mínimo de veinte otras modalidades metabólicas no accesibles a ningún animal o planta, a menos de que las adquiera por medio de la simbiosis. Algunas bacterias respiran arsénico o azufre en lugar de oxígeno. Otras brillan en la oscuridad gracias a la producción de «luz fría», fruto de la reacción luminiscente entre la luciferina y la luciferasa. Otras bacterias medran en comunidades pestilentes, donde sus miembros nadadores transforman los iones metálicos de las aguas en precipitados metálicos tales como óxidos de manganeso o de hierro, o incluso oro. Algunas bacterias azuladas o verdosas fotosintetizan y expelen oxígeno cuando están expuestas a la luz, mientras que cuando se hallan en la oscuridad aspiran oxígeno como los animales y los hongos. Otras utilizan la luz del sol para fotosintetizar de formas que o bien impiden la liberación, o bien la absorción de oxígeno. Otras bacterias, denominadas metanógenas, convierten el C02 y el hidrógeno en gas de los pantanos. En total ausencia de gas oxígeno y de alimento, producen metano (CH4), exactamente el mismo gas que empleamos en nuestros quemadores de «gas natural». El metano desciende hasta las fisuras donde otra bacteria (metilótrofa) lo quema. Las metilótrofas respiran metano y sacan de él la energía necesaria para medrar y crecer. Otras aún viven y se desarrollan en las profundidades marinas, gracias a reacciones químicas en miniatura basadas en la luz, que los científicos más avezados aún no han conseguido ni explicar ni imitar.
Comprender los entresijos de la evolución requiere comprender el verdadero espectro de las posibilidades de la vida. En la flexibilidad, el virtuosismo cosmopolita y las sutiles estrategias de las bacterias, podemos comenzar a apreciar la complejidad de la evolución en su verdadero esplendor. La mayor parte de las bacterias rechazan la compañía de los humanos. Mucho antes de que el primer animal comenzara a deambular por la Tierra, ellas ya se reproducían, exploraban, conquistaban, intercambiaban genes, se deleitaban en sus sorprendentes formas de encuentro sexual y erosionaban rocas. Sin duda han orbitado alrededor de la Luna y han alunizado sobre su suelo muchas más bacterias que seres humanos. Que las bacterias se dupliquen, transfieran genes, digieran y ganen o pierdan genes de formas muy diversas, constituye a no dudar un aspecto esencial en la saga de la evolución. La velocidad, el volumen y la antigüedad de las actividades bacterianas de intercambio de genes subyacen en la evolución de todas las demás formas de vida sobre el planeta. La Humanidad es una mota de polvo en la biosfera, un experimento sin acabar de verificar, susceptible de extinguirse por autoconsunción. La célula nucleada constituyó el resultado más espectacular de las maquinaciones bacterianas. A diferencia de la Humanidad, ella sí que ha sido plenamente verificada.
§. Incorporaciones: donde la especiación comienza
Constituyendo equipos que se integraban para formar las primeras células nucleadas, las bacterias iniciaron el proceso de especiación. Algunas de aquellas células nuevas se convirtieron en descendientes protistas, evolucionando nuevos paquetes de capacidades que les permitieron abandonar el reino bacteriano. Fue ahí donde se iniciaría el proceso de especiación, en el mundo del Proterozoico inferior, hace unos 2.000 millones de años.
Los protoctistas ampliaron el mundo de las bacterias. Del mismo modo que los animales formadores de embriones que les sucedieron, desarrollaron nuevas estrategias tróficas y sociales. Fue ahí, en el mundo protoctista, donde los primeros ingenieros y diseñadores crearon estructuras tales como delicadas cajitas de sílice o plataformas de caza de carbonato cálcico. Otros inventaron la agricultura. Los foraminíferos unicelulares, representativos de unas 60.000 especies distintas y que incluyen células de tamaño relativamente grande, atrapaban y cultivaban algas que expelían por las ranuras de su cuerpo y almacenaban en los pesebres de sus cáscaras. Esas algas, bien entrenadas, absorbían durante el día la luz solar y producían alimento a partir de ella. Luego, abrigadas durante la noche en los orificios de la cáscara de su anfitrión, respiraban el oxígeno que habían producido a lo largo del día. Algunos parientes lejanos de estos agricultores construyeron nuevas cáscaras a partir de mezclas multicolores. Estos «aglutinadores» pegaban piedrecillas a su cuerpo. Otros foraminíferos incluso reúnen minerales diversos para construir con ellos torres de vigilancia, por las que ascienden para cazar animales mucho mayores que ellos, tales como rotíferos y crustáceos. Los protoctistas incluyen grandes «ambigüedades acuáticas», organismos que no son ni animales ni plantas como el kelp, un fotosintetizador de cien metros de longitud que sigue presente en nuestros días. El registro fósil de los foraminíferos y las diatomeas es inmenso, incluso si lo comparamos con el de los animales marinos con cáscara, tales como los bivalvos y los trilobites. Sin embargo, escasean las descripciones de protoctistas debido a la carencia de paleontólogos y protistólogos vivos que se ocupen de ello. A no dudar, podríamos aprender mucho de sus tecnologías en miniatura y de sus asociaciones simbióticas si nos lo propusiéramos.
§. Ecuaciones celulares y definiciones de especies
Hace unos 450 millones de años, los hongos y las plantas se unieron a las bacterias, a los protoctistas y a los animales. Mientras que las plantas pertenecen al mundo de lo fácilmente visible, la mayoría de hongos, como las levaduras y los mohos, encajan en la descripción de los microbios. A pesar de que las setas y algunos mohos grandes y complejos son fácilmente reconocibles para el ojo del profano, incluso en su caso —al igual que sucede con la totalidad de los microbios— se les puede observar y comprender mejor con la ayuda del microscopio. A simple vista, la mayoría de hongos no son más que un polvillo blancuzco. Los micólogos profesionales y los aficionados ocasionales —cuyo número disminuye año tras año— son en la práctica las únicas personas capaces de distinguir una especie de otra.

En resumen, los microbios (bacterias procariotas, protoctistas eucariotas y hongos) evolucionaron para actuar de formas que van mucho más allá de las prácticas y los estilos de vida relativamente uniformes de los animales y las plantas. Para comprender las artimañas del proceso evolutivo de animales y plantas necesitaremos profundizar en el conocimiento del microcosmos y de los talentos, las necesidades, los exudados y las peculiaridades de género y reproducción de sus miembros. Los microbios tienen genomas completos de capacidad única. Son ellos, y no los genes egoístas ni los mamíferos combativos, los verdaderos motores del cambio evolutivo.
En la tabla 6.1 se incluyen las ecuaciones abreviadas de los sistemas genómicos integrados de la mayoría de grandes formas de vida complejas.
Capítulo 7
La historia de lo heredable
Bajo estrés, distintas clases de individuos, con orígenes y capacidades muy diversas, tienden a asociarse físicamente. Frente a un estrés continuado y predecible, frecuentemente cíclico o estacional, estas asociaciones se vuelven íntimas y se extienden más allá de un encuentro ocasional. Para convertirse en significativos para el proceso evolutivo, los anteriores extraños deben interactuar con la suficiente frecuencia como para llegar a formar una relación única y estable, que acabe por desembocar en una relación permanente, o al menos firmemente estacional. Para expresarlo de forma resumida, en los casos importantes para la evolución, las asociaciones condujeron a simbiosis que desembocaron en nuevas clases de individuos, formados por simbiogénesis. En cualquier momento esas asociaciones pueden disolverse, sus miembros pueden cambiar e incluso destruirse entre sí, o simplemente perder a su simbionte. Los resultados de la interacción de organismos vivos muy distintos no son plenamente predecibles. En ese contexto, los términos de “coste” y “beneficio” no resultan demasiado útiles. Las asociaciones pueden hacerse tan íntimas que se llegue a producir literalmente una incorporación. De la asociación casual y de su inevitable cambio metabólico (donde los residuos de uno de los organismos, en un sistema termodinámico abierto, se convierten en alimento, protección o lubricante para el otro) surgen entonces nuevos cuerpos mutuamente incorporados. Los organismo otrora independientes pasan a ser reconocidos como componentes de nuevas fusiones, sin perder del todo los vestigios de su anterior estado autónomo. Los cloroplastos verdes de las hojas siguen subdividiéndose. Los bordes batientes de las espiroquetas siguen nadando dentro de la Mixotricha, sólo que no se escapan nadando. Tras dilatados períodos de vida complementaria, asociación íntima o dependencia metabólica, los extraños compañeros de cama terminan a menudo por fundir sus sistemas genéticos. Es la etapa final consistente en la adquisición de genomas, la clave de la «herencia de genomas adquiridos». Los socios otrora separables y distintamente nombrados se convierten ahora en una nueva y única entidad, en un individuo nuevo situado en un nivel superior de tamaño y complejidad. Están documentadas numerosas fusiones de esta índole en la totalidad de los cinco reinos de la vida.
Las asociaciones que más nos interesan para lo que aquí nos ocupa son aquellas que espolearon innovaciones discontinuas y evidentes en la evolución, tan notables que las huellas de su historial quedaron claramente impresas en los registros fósiles. Estos ejemplos sobresalientes incluyen por igual a los animales de los arrecifes coralinos y a sus algas, a los bivalvos fotosintéticos, a los gusanos tubiformes oxidadores de azufre, de dos metros de largo, de los abismos oceánicos, a las termitas urbanizadas y a las vacas lecheras.
Las plantas jaspeadas llaman nuestra atención. Nuestra mirada se siente atraída por las manchas de color, las franjas o los dibujos sobre hojas, frutos, flores o tallos. Las razones por las que las plantas exhiben estas decoraciones son diversas, pero entre ellas se cuenta sin duda la infección vírica. En principio, los virus son demasiado pequeños como para poder funcionar por sí solos, pero sus genomas penetran por igual en las células de plantas, animales, protoctistas, bacterias y hongos, para aprovecharse del metabolismo de sus anfitriones. Manchas imperceptibles pueden convertirse en bellas decoraciones cuando virus específicos son introducidos en plantas nuevas mediante injertos de la rama o el tallo infectados procedentes de otra planta. Por ejemplo, los virus en los tulipanes híbridos o en determinada especie de Abutilón (Abutilón pictus) causan hermosas franjas o hermosas manchas de color crema y amarillo brillante. Los virus no perjudican a la planta en absoluto, y la gente selecciona y propaga esas plantas por su valor estético. He aquí otro ejemplo de adquisición de genomas como fuente de variación heredable, provocada dentro de una especie por un virus y mantenida en la población por los jardineros (Marcotrigiano 1999, figura 5).
§. Presentamos a los escarabajos. El gorgojo de Heddi
El gorgojo, pesadilla del hombre neolítico a medida que sus huertos de grano se iban extendiendo, sigue con nosotros hoy en día. Ese escarabajo minúsculo, del que abundan las variedades, vive enteramente entre las gramíneas y las poáceas, miembros ambas de la familia de las hierbas. Las segundas constituyen la herencia de la expansión global de nuestras especies alimenticias del trigo, el maíz, la cebada y la avena. El mejor estudiado de estos insectos es el Sitophilus oryzae, conocido como gorgojo del arroz pero perfectamente capaz de destruir el grano almacenado de otras clases. Todo su ciclo vital tiene lugar dentro de un grano de arroz o de trigo. Hay quienes opinan que ese escarabajo evoluciona con nosotros. Aparentemente ello se debe a que descarta los genes que no necesita a medida que le proporcionamos los productos genéticos que ya no tiene que producir por sí mismo.
Durante setenta años, la presencia de esos escarabajos de tejido bacteriano —otrora denominados micetomas porque se creía que su origen era fúngico— ha sido objeto de investigación. Un órgano larval, situado en el ápice del ovario de la hembra y denominado bacterioma, está repleto de bacteriocitos. Estas células hinchadas del escarabajo albergan una enterobacteria que presenta una similitud genética del 95 por ciento con la bacteria común del colon, la Escherichia coli. Detectable tan sólo en tres partes del organismo del insecto —en el bacterioma larval, tanto en hembras como en machos, en el bacterioma del ovario de la hembra y en los huevos o células germinales de la misma—, esta bacteria, al igual que nuestras mitocondrias, es heredada por parte de madre. Ello implica que los descendientes del gorgojo, tanto si son machos como si son hembras, hereden de sus madres las células que formarán el bacterioma. El esperma no tiene nada que ver en todo ello. ¿Qué proporcionan esas bacterias? Al menos una parte de la respuesta, extraída de la Naturaleza gracias al trabajo ingente de numerosos investigadores, consiste en vitamina B. El genoma bacteriano está repleto de genes que codifican estos suplementos cruciales en la nutrición, entre los que se cuenta la riboflavina, el ácido pantoténico y la biotina. Cuando se eliminan esas bacterias de los tejidos del escarabajo, ya sea con antibióticos o por otros medios, la tasa de crecimiento del gorgojo se ve marcadamente reducida. Los escarabajos privados de bacterias maduran más tarde que los que mantienen su dotación de simbiontes. Más importante aún, habida cuenta de que su suministro de energía mitocóndrica se ve alterado, los gorgojos privados de sus simbiontes bacterianos no pueden volar, metáfora muy indicada para la importancia de la simbiogénesis en la Naturaleza.
Expertos en el gorgojo, como Abdelazziz Heddi y su mentor, el profesor Paul Nardon, del Institute des Sciences Appliquées de Lyon, denominan EPSO (Endosimbionte Primario del Sitophilus oryzae) [12] a esta bacteria altamente integrada. Desde China o Guadalupe a su huerto de Lyon o a Italia, donde Umberto Pierantoni (1876-1959) lo descubriera por primera vez, todas las ramas del gorgojo Sitophilus oryzae contienen la misma numerosa población de esa bacteria, siguiendo un mismo patrón. Es probable que la adquisición e integración de la bacteria simbiótica en el metabolismo del gorgojo coincida con el origen de la especie oryzae.
Pero eso no es todo. Heddi y sus colaboradores encontraron a finales de los 90 otra clase de bacterias asociadas con el gorgojo. De entre el par de docenas de linajes en todo el mundo, que representan a tres especies de Sitophilus (oryzae, granarías y zeamais), el 57 por ciento demostraron albergar en sus tejidos a una bacteria más. A diferencia de la EPSO, cuando se «curó» (por medio de antibióticos) a los gorgojos de esta segunda bacteria, no apareció ningún efecto en la fisiología de los escarabajos, ni en su metabolismo del nitrógeno ni en su capacidad para volar. Más bien parecía que esa bacteria, parecida al linaje Wolbachia, fuera mucho menos regular en lo relativo a sus costumbres. El nuevo simbionte bacteriano se encuentra en número variable en todo el cuerpo del insecto, salvo una excepción: abunda en los tejidos germinales (sexuales), particularmente allí donde puede interactuar con núcleos de esperma en desarrollo. Algunos investigadores sospechan que la nueva bacteria se adhiere a las peculiares proteínas fijadoras de cromatina de los tejidos productores de esperma. Sean cuales fueren los detalles de su acción, esta bacteria incide sobre la fertilidad de esos prolíficos comedores de granos. Al aparear a machos dotados de la bacteria parecida a la Wolbachia con hembras que carecían de ella, la fertilidad decreció espectacularmente. El cruce inverso, es decir, entre hembras con la bacteria y machos sin ella, produjo también un descenso en la fertilidad, aunque menos notorio. Para gran ventaja de la bacteria oportunista, el número de descendientes era el máximo cuando ésta estaba presente en ambos progenitores. El efecto de la simbiosis consiste, por consiguiente, en promover el aislamiento reproductivo. Los apareamientos entre gorgojos no infectados son por supuesto fértiles, y aquéllos producidos entre consortes infectados también lo son. Sin embargo, los apareamientos mixtos lo son menos. El aislamiento reproductivo constituye, por supuesto, uno de los elementos clave de la especiación.
El concepto de que el aislamiento reproductivo —y por consiguiente la especiación incipiente— pueda ser inducido por la presencia de simbiontes microbianos no es nuevo en la bibliografía biológica. La idea fue bien argumentada por Theodore Dobzhansky y sus colegas, que se dedicaron al estudio de la Drosophila en poblaciones en cautiverio. Aparearon moscas de la fruta que habían sido sometidas a diferentes temperaturas (frío y calor) durante un par de años. Los apareamientos anteriores habían sido plenamente fértiles, pero al aparear luego a moscas criadas en el frío con otras criadas en el calor, los resultados fueron menos fértiles. La causa probable de esta merma en la fertilidad era la presencia de micoplasmas (bacterias sin paredes celulares) en las condiciones de frío, así como la pérdida de este invasor de tejidos a temperaturas superiores en las poblaciones incubadas con calor. Nardon, Heddi y muchos otros autores han documentado esta misma observación: si macho y hembra de la misma especie llevan en sus tejidos las mismas bacterias, su apareamiento produce descendencia fértil normal. Los problemas comienzan cuando un miembro de la pareja las lleva y el otro no.
Dobzhansky y los demás, aun estando en lo cierto, nunca fueron suficientemente explícitos. Tomaron nota de la presencia y la ausencia de la bacteria, así como de sus efectos sobre la fertilidad, pero nunca llegaron a elevar esta observación a la categoría de mecanismo general de promoción de la especiación. Cuando los micoplasmas o las proteobacterias eran adquiridos por uno de los dos géneros de insectos, impidiendo la fertilidad normal a menos que el otro insecto incorporara también el nuevo microbio, las consecuencias eran el «aislamiento reproductivo» y la especiación. La atracción de las bacterias al tejido reproductivo tanto en hembras como en machos, particularmente en los insectos, fue abundantemente documentada por Paul Buchner (1886-1978) y sus sucesores.
De nuevo definimos a las especies como sigue:
Dos seres vivos pertenecen a la misma especie cuando el contenido y el número de genomas integrados, previamente independientes, que los constituyen, son los mismos. El concepto de especie de E. Mayr en 1948, que afirma que los organismos pueden ser asignados a una misma especie cuando, por naturaleza propia, se aparean y producen descendencia fértil, se convierte así en un ejemplo de nuestra regla general. El concepto de Mayr resulta particularmente aplicable a los animales, que ingieren cada día bacterias susceptibles de pasar de su intestino a sus gónadas o a otros tejidos grasos reproductivos. Esos minúsculos forasteros pueden nadar por sus tejidos, hallar un nicho, permanecer en él e influir en el futuro de lo que podría ser una nueva especie, con un conjunto enteramente nuevo de genes útiles: el genoma adquirido de la antes talentosa, pero ahora perezosa, bacteria incorporada.§. Diálogo microcósmico: integración de socios
La pregunta de Darwin sobre cómo se originan las especies puede ser reformulada como sigue: « ¿qué pasa de progenitores a descendientes, que pueda ser detectado como novedad evolutiva?» Una respuesta directa consiste en: «poblaciones y comunidades de microbios». Recordemos que las poblaciones son grupos de individuos del mismo tipo que viven en el mismo lugar al mismo tiempo. Sin embargo, lo que prevalece en la Naturaleza son las comunidades, es decir, las agrupaciones de poblaciones de organismos diversos. Cuando miembros de una comunidad —diferentes formas de vida conviviendo (o meramente sobreviviendo) en un mismo lugar al mismo tiempo— se fusionan y transfieren genes entre sí, evolucionan «individuos» nuevos y más complejos. Invariablemente, los biólogos asignan nuevos nombres de especie a esos individuos novedosos, mayores y más complejos.
Las historias acerca de cómo los microbios tienden a unirse entre sí, así como de las múltiples interacciones entre ellos y con organismos mayores, han sido contadas varias veces en los lenguajes especializados de las ciencias. Inevitablemente, esas historias son poco conocidas, debido en parte a la gran fragmentación de la propia ciencia. Incluso aquellos de entre nosotros que comprendemos cuánto se conoce ya realmente acerca de los orígenes de las especies, nos vemos limitados a trabajar sobre nuestros minúsculos descubrimientos, habitualmente sobre una sola especie cada vez. En otros tiempos, los departamentos académicos de biología reunían a zoólogos y botánicos. Las plantas y los animales se estudiaban conjuntamente. Relegado a los resquicios de las instituciones de las artes liberales, el estudio de los microbios quedaba restringido a sus propios departamentos, denominados de microbiología, todos los cuales residían en escuelas de medicina o de agricultura. El objetivo principal del conocimiento sobre los microbios —vistos como agentes patológicos o como contaminantes alimentarios— consistía en descubrir el mejor modo de destruirlos.
Desde los años 80, numerosos departamentos de ciencia biológica se han dividido aún más entre biología molecular frente a biología organísmica, tendencia que no hace más que exacerbar la incomprensión. La información relevante sobre los orígenes de las especies se halla dispersa en más de una docena de campos de estudio distintos, dotado cada uno de los cuales de su propio lenguaje o lenguajes esotéricos. A veces, organismos idénticos se ven repartidos entre disciplinas diferentes. Las cianobacterias, por ejemplo, se estudian dentro de la ficología (o algalogía, una rama de la botánica) y no en la bacteriología, simplemente porque antaño fueron erróneamente denominadas «algas verdeazuladas». Para el descifrado del origen de las especies son relevantes la bioquímica, la biología celular, la geología, la zoología invertebrada, el metabolismo, la evolución molecular, la ecología microbiana, la nutrición, la paleontología, la protistología, la geología sedimental y la virología. La mayoría de esos campos, verdaderas cajas negras tanto para el gran público como para los graduados, siguen siendo un misterio, incluso para muchos de los científicos que practican la biología evolutiva en la actualidad.
Es frecuente que muchos de los famosos pero mal encaminados seguidores de Darwin, interpreten erróneamente incluso las partes de la ciencia que conocen bien. Las revelaciones de gran parte de la ciencia más allá de su siglo, ampliadas por la biología molecular y la paleontología, son completamente coherentes con la intuición de Darwin. Sin embargo, estas revelaciones demuestran que la lujuriante diversidad viva que nos rodea no evolucionó de forma gradual, como no se cansan de repetimos quienes se dedican al estudio del registro fósil. Son muy escasos los restos en rocas sedimentarias que acrediten la existencia real de una serie de pequeños pasos, que conecten gradualmente a una especie con sus descendientes. Las «huellas de biosferas pasadas», por utilizar la frase inmortal de Vladimir Vernadsky, proclaman exactamente lo contrario. El equilibrio puntuado está ahí bien patente para quien lo quiera ver. El registro discontinuo de la vida pasada demuestra claramente que la transición de una especie a otra ocurre en saltos discretos. En trilobites, caracoles, esporas, peces dipnoos, tiburones y bivalvos, abundan las pruebas de cambio puntuado. A pesar de la queja del propio Darwin acerca de lo incompleto del registro y su postulado de un «paso» desde una a otra forma de vida, por ejemplo en las palomas, los percebes o los perros, la vida animal y vegetal ha evolucionado en pasos del tamaño del genoma microbiano. Lo que aparece como «magia», «complejidad irreductible» o «gran plan» —la imagen de la retina del ojo del vertebrado, el vuelo de abejorro, el canto de la ballena jorobada que cruza el océano o el de los Tres Tenores (Luciano Pavarotti, José Carreras y Plácido Domingo)— no es ni más ni menos que el legado de interacciones repetidas. El diálogo metabólico y la proximidad física condujeron a la incorporación, a la acomodación y al reajuste. Los organismos conocidos no son única y simplemente individuos, sino que surgen del submundo microbiano.
¿Cómo hacen entonces los organismos independientes y separados para fusionarse y formar individuos nuevos? ¿Cómo logran estos individuos retener y perpetuar sus seres compuestos? ¿A qué nos referimos cuando hablamos de «adquisición e integración» de genomas? En unos pocos casos específicos, las respuestas a estas preguntas son conocidas porque podemos encontrar en la Naturaleza a los organismos independientes de distintas clases, podemos reconocer formas intermedias (más o menos integradas) y con todo ello nos aventuramos a reconstruir la historia del organismo compuesto.
Ivan Emmanuel Wallin (1890-1967), profesor de anatomía en la Escuela de Medicina de la Universidad de Colorado, escribió en 1927, cuando era aún bastante joven y vivía en Nueva York, Symbionticism and the Origin of Species. La de Wallin era por aquel entonces una voz solitaria que proclamaba la importancia capital de la simbiosis en el proceso evolutivo. Wallin inventó su propio nombre para la simbiogénesis microbiana, que denominó formación de «complejos microsimbióticos». Apodó «simbionticismo» al proceso de integración simbiótica en la evolución, sin percatarse de que en Europa y desde 1860 se había estado desarrollando una extensa bibliografía sobre el mismo concepto. Nunca supo, por ejemplo, del trabajo de Konstantin Merezhkousky o de Andrey Faminstyn. La bibliografía europea, publicada en ruso, francés y alemán, escapaba a Wallin porque, al igual que la mayor parte de los científicos estadounidenses, sólo sabía inglés. Desde finales del siglo XIX, en el mundo científico europeo la adquisición y herencia de simbiontes microbianos se había venido denominando «simbiogénesis». El darwinismo en general era desdeñado. Se decía que, desde un principio, la selección natural no podía producir «buenos» cambios evolutivos. La simbiogénesis era considerada como crucial para la generación de novedad biológica. La bibliografía rusa, interpretada por el historiador de la ciencia Liya N. Khakhina, no estuvo disponible en inglés hasta el año 1922. Fueron necesarias dos generaciones de académicos para resumir la gran bibliografía de los botánicos rusos. Parece hoy como si esta bibliografía fuera ignorada por esta misma razón. La literatura antigua escrita por botánicos rusos carece de atractivo para el mercado anglófono. No obstante, los científicos rusos (principalmente Faminstyn, Merezhkousky y Kozo-Polyansky, cada uno de ellos con distinto énfasis) presentaron la misma tesis que Wallin: las simbiosis y sus consecuencias, particularmente la simbiosis microbiana, generan cambio evolutivo.
Wallin desarrolló primero una teoría de «prototaxis» para responder a su pregunta acerca de cómo estas simbiosis íntimamente integradas pudieron llegar a iniciarse. Como todos quienes nos dedicamos a este campo de la ciencia, Wallin se percató de que todos los organismos son producto de su historia altamente específica. La prototaxis, según Wallin, precedía a la simbiosis en cualquier asociación de más de un único tipo de vida. Con el término “prototaxis” Wallin se refería a la tendencia innata de una clase de célula u organismo a responder de un modo específico a otra clase de organismo (Wallin 1927, pág. 8).
Los ejemplos incluyen la tendencia del ratón a huir del gato, del tiburón a tragarse el pez, del conejo a comer lechuga o de la mosca a poner sus huevos en el tejido muscular sanguinolento del jabalí recién abatido. Como demuestran estos ejemplos, la prototaxis puede ser tanto positiva como negativa, tanto asociativa como disociativa. Lo sabemos bien por experiencia propia. Instintivamente huimos del enjambre de abejas y nadamos hacia el macizo de lirios acuáticos. La prototaxis está en el inicio de cualquier caso de simbiogénesis.
La fotosíntesis, el fenómeno químico natural que transforma la luz y el dióxido de carbono del aire en energía y alimento, es enteramente bacteriana. Todos los organismos fotosintéticos, ya sean bacterias o descendientes de bacterias, extraen compuestos orgánicos a partir del botín de su luz cosechada. El comportamiento prototáctico de numerosas formas de vida no fotosintéticas hacia los fotosintetizadores resulta fácilmente comprensible. La motilidad, ya sea nadando, gateando, resbalando o arrastrándose, sirve para garantizar que los seres hambrientos, incapaces de realizar por sí mismos la fotosíntesis, accedan a las zonas bien soleadas imperativas para los organismos fotosintetizadores. La tendencia prototáctica de los «heterótrofos» a absorber los productos de la fotosíntesis, o bien a ingerir a los propios organismos fotosintetizadores, ha conducido a la explosión de prósperas comunidades de comedores en las zonas soleadas de las superficies de agua dulce y salada. Los organismos secundariamente fotosintéticos —aquellos que adquirieron la fotosíntesis por haber ingerido las bacterias adecuadas sin lograr sin embargo digerirlas— incluyen a la totalidad de eucariotas fotosintéticos. Ningún alga o planta evolucionó nunca la fotosíntesis por sí sola. Todas comparten algún antepasado, reciente o remoto, que ingirió pero que no digirió algún fotosintetizador bacteriano verde, rojo o verde-azulado. En este caso, la prototaxis es la tendencia hacia el hambre por parte del comedor y hacia la resistencia a la digestión por la del comido. En resumen, el hambre a la luz del sol y la resistencia a ser digerido han conducido, una y otra vez, a organismos fotosintéticos pigmentados permanentes: algas, líquenes, plantas, lombrices verdes, corales marrones, hidras verdes y bivalvos gigantes (Tridacna) —cuyas inmensas cáscaras permanecen abiertas para enfocar la luz hacia los cloroplastos de sus algas simbióticas— constituyen tan sólo unos pocos de los muchos ejemplos de los que depende nuestra vida. Las prototaxis, estas tendencias orgánicas, pueden constituir las versiones iniciales de esa clase de propósito que, en nuestro propio caso, denominamos «elección consciente». Sin embargo, todos los seres tienen una relación con el tiempo que es más compleja que la simple duración: están orientados, por sus acciones asociativas o disociativas, hacia las consecuencias futuras.
Los socios simbióticos se integran a diferentes niveles. El primero y más superficial de ellos consiste en el comportamiento. Para que cualquier asociación permanente pueda iniciarse, es indispensable que ambos candidatos potenciales se encuentren en el mismo lugar al mismo tiempo. ¿Qué les lleva a ese charco iluminado, a la copa de ese árbol, a la caverna sombría o a perderse en la niebla? El pasado y la contingencia determinan el comportamiento inmediato de todo organismo. Algunas asociaciones simbióticas estables están integradas únicamente al nivel del comportamiento. No obstante, otras muchas alcanzan niveles de integración más íntimos, desde lo conductual a lo metabólico. Frecuentemente el producto metabólico, el exudado o el residuo de uno de los miembros de la asociación se convierte en alimento para el otro. Probablemente todos los animales verdes que han sido estudiados (tales como la lombriz plana Convoluta roscoffensis, o la Hydra viridis de los estanques), así como todos los líquenes, estén integrados a este nivel. Los «fotosintatos», es decir, los azúcares u otros productos de fotosíntesis elaborados por el socio verde, pasan a ser engullidos como alimento por el otro socio. Puede suceder que el fotosintato sea vertido directamente en el cuerpo del recipiente. Éste, a su vez, inevitablemente orina, defeca o produce sus propios residuos a su manera, a menudo ricos en nitrógeno, que se convierten también en un nutriente necesario para el fotosintetizador. Esta clase de integración al nivel metabólico es bastante corriente. Cuando los miembros de la asociación tienen una dilatada historia en común, su integración no resulta fácilmente reversible y sus individualidades se diluyen. El parásito de la malaria (Plasmodium vivax) necesita conseguir su alimento del torrente sanguíneo de su víctima. En este caso, el protista y el mamífero (Homo sapiens) están claramente integrados al nivel metabólico. Los líquenes lo están también claramente. Los productos de la fotosíntesis fluyen desde el socio verde o verdeazulado hacia el socio translúcido. Raramente se percibe que, en las asociaciones íntimas, los metabolitos fluyen en ambas direcciones. El socio animal o fúngico también libera materiales para el fotosintetizador. Las simbiosis son intercambios de doble sentido. La amabilidad del extraño, el flujo de presentes metabólicos, hace que resulte menos extraño hasta que, finalmente, se convierte en parte integrante de un ser biológico único y co-dependiente. Ver Brodo et al., 2001.
Las asociaciones metabólicas pueden integrar aún más estrechamente. El siguiente nivel de asociación se conoce con el nombre de «asociación productora de genes». En ella, las proteínas (o incluso moléculas de ARN) de uno de los miembros son necesarias para el funcionamiento del otro. En las plantas de alubias y guisantes encontramos un excelente ejemplo de esta clase de integración. Si arrancas un trébol, una arveja o una planta de judías, verás en sus raíces unas pequeñas protuberancias rosáceas. Se trata de nódulos fijadores de nitrógeno, en cuyo interior medra determinado tipo de bacteria. Otrora bacterias nadadoras con forma de bastoncito, todas ellas han acabado por convertirse en «bacteroides» hinchados. Estos bacteroides sobredimensionados, llenos de agujeros, ya no pueden dividirse ni crecer. El color rosáceo de los nódulos resulta de una proteína denominada hemoglobina, la misma clase de proteína —aunque diferente en los detalles— que hace que nuestra sangre sea roja. En nuestra sangre, la hemoglobina transporta el oxígeno de los pulmones a los tejidos que precisan de él. En el caso de las plantas de judías y guisantes, la hemoglobina se ocupa de alejar el oxígeno de donde están las enzimas fijadoras de nitrógeno, ya que este gas las envenenaría. Desde el punto de vista de la simbiosis entre la planta y las bacterias fijadoras de nitrógeno, el punto crucial estriba en que la molécula de hemoglobina es en sí misma quimérica. Tanto en las plantas como en los seres humanos, las hemoglobinas tienen dos componentes: uno es la proteína (la globina), mientras que el otro es la parte que le da color. Este segundo componente, una molécula no proteinácea mucho más pequeña, recibe el nombre de heme. En las plantas con nódulos, la heme es producida por el socio bacteriano, mientras que la proteína globina es producto de la planta. He aquí una historia de integración de productos genéticos en una simbiosis. El producto final —la molécula de hemoglobina perfectamente constituida— no llegaría a existir sin las respectivas contribuciones de la bacteria y de la planta.
El grado máximo de intimidad entre socios implica la integración al nivel genético. Cuando un gen de un organismo se incorpora y permanece entre los genes de otro —por ejemplo, cuando un gen pasa de una bacteria viva al núcleo celular de una planta—, la integración es completa. No se conoce mayor grado de intimidad que la adquisición permanente de los genes del socio. Para cuando ocurre esta clase de intimidad, lo más probable es que ya estén presentes las integraciones a los niveles de comportamiento, metabolismo y productos genéticos. Cuando las asociaciones perduran, se vuelven íntimas y comparten historias dilatadas. Asimismo, los factores que impiden su disolución son numerosos. Lo más frecuente es que los miembros de semejante asociación no puedan ya volver jamás a su estado anterior.
Si le das la vuelta a una estrella de mar de la costa de California verás multitud de pequeñas lombrices negras alrededor de su boca. A través de este orificio, el animal ingiere gran cantidad de alimento, pero las lombrices no son ni dañadas ni engullidas. Si separas a estas lombrices de la boca de la estrella, rápidamente se volverán a colocar a su alrededor. Las estrellas de mar son agresivas. Las lombrices se alimentan de restos y fragmentos de comida, justo en la misma boca del lobo. Esta asociación simbiótica entre lombriz y estrella de mar es exclusivamente conductual. Dos sustancias producidas por la estrella afectan a las lombrices. La primera (fácilmente destructible por el calor y, por consiguiente, probablemente una proteína) hace alejarse rápidamente a las lombrices. La segunda (un compuesto estable ante el calor) las atrae hacia los aledaños de la boca de la estrella. Combinados el uno con el otro, ambos compuestos logran mantener a las lombrices dentro de su ámbito de alimentación. Debe estar también presente alguna otra complejidad química, puesto que la estrella de mar no se come a esas lombrices, pero sí a sus parientes y a todo lo que caiga al alcance de su temible boca. Probablemente algún compuesto inhibidor previene la ingestión de estas vulnerables lombrices negras en concreto que, con su frenético picoteo de restos de comida, mantienen el área de la boca escrupulosamente limpia, proporcionando a su anfitrión una versión natural de higiene bucal.
Asociaciones laxas, de índole conductual, como la que acabamos de ver, pueden llegar a volverse indefinidamente estables. Nos encontramos con esta clase de asociación en determinadas situaciones, tales como inviernos fríos que provocan la formación alternativa de charcas húmedas y suelos helados y secos, en las que organismos no asociados dejan más descendencia por separado que la que dejarían juntos. En otra época del año, las circunstancias favorecen la asociación simbiótica. El resultado consiste en que la simbiosis conductual persiste, pero no crece en intimidad. La integración completa e irreversible de dos seres diferentes para formar otro nuevo ocurrirá cuando, en cualquier situación, los organismos físicamente asociados producen más descendientes que si no lo estuvieran. Por ejemplo, los centenares de mitocondrias de cada una de nuestras células nunca las abandonan. ¿Por qué? Porque el universo de los tejidos animales está repleto de oxígeno y requiere que este gas fluya continuamente a cada célula. En el Proterozoico, rico en oxígeno, las células que retuvieron a las mitocondrias de forma permanente debieron ser «seleccionadas naturalmente» sobre aquellas que dejaron retomar a las mitocondrias a su mundo bacteriano. Por consiguiente, es necesario preguntarse en todos los casos cómo están integrados los miembros de la asociación, si siempre lo están, y qué condiciones ambientales influyen sobre su integración. Sustituir esta clase de detalles de flujo metabólico y transferencia de productos genéticos entre organismos íntimos, precedentemente extraños, por términos neodarwinianos tales como “cooperación”, “coste” o “beneficio”, es completamente absurdo y representativo de «la falacia de la concreción mal atribuida». Semejante terminología impide la verdadera comprensión del pasado evolutivo inevitablemente rico y complejo del mundo simbiótico que creó a los animales, a las plantas y a sus compañeros de viaje nucleados. Como dulce azucarado, que nos sacia temporalmente pero que al mismo tiempo nos impide alimentamos correctamente, el neodarwinismo sacia la curiosidad intelectual con abstracciones ajenas a los detalles reales, sean éstos metabólicos, bioquímicos ecológicos o pertenecientes a la historia natural.
§. Agricultores fúngicos
Desde el Mesozoico, desde hace probablemente 100 millones de años, las termitas han venido viviendo en los troncos de árboles caídos, entre cuya madera se han alimentado y han construido sus nidos. Todas las termitas son insectos, clasificadas dentro del filum animal de los artrópodos y pertenecientes al orden de los isópteros (iso = igual; ptera = alas).

Figura 7.1 Gamba roja limpiadora Lysmata grabhami en la boca de la morena verde Gymnothorax funebris.
Todas las demás termitas, con mucho la mayor parte del taxón, se consideran «superiores», es decir, evolucionadas con posterioridad, lo cual no significa más que los antepasados de estas termitas abandonaron a sus fascinantes ninfas microbianas simbióticas intestinales, digestoras de madera, evolucionando estrategias de búsqueda de alimento más eclécticas. Algunas de estas termitas llamadas superiores ensancharon sus gustos, disfrutando actualmente de ricas dietas orgánicas. Otras siguen totalmente restringidas al antiguo régimen de madera. La lignina y la celulosa son sus únicas fuentes de carbono. Sobrevivieron a su pasado paleolítico, pero siguen alimentándose exclusivamente de madera de todas clases. Estas termitas inventaron una agricultura con la que cultivan un socio digestor de madera, un hongo (o tal vez varios hongos similares) al que se puso el lógico nombre de Termitomyces. Las termitas socialmente «superiores» domesticaron estos hongos, que siembran y recogen.
Los micólogos experimentan cierto sonrojo y tal vez deberían sentirse genuinamente incómodos, ante su torpeza en lograr cultivos puros de Termitomyces, si comparamos sus resultados con la habilidad demostrada por las termitas agricultoras, dotadas de mucha más experiencia. Las activas termitas superiores labran y preparan el suelo, lo fertilizan, podan los brotes malsanos de hongos denominados hifas, y limpian el cultivo de malezas tales como hongos no deseados u otros restos. Finalmente, al final de la temporada recogen su cosecha. ¡Ni siquiera el más experimentado micólogo logra aproximarse a la habilidad demostrada por las termitas en el cultivo de Termitomyces!
«El efecto de la poda y el cultivo se entiende mejor al comparar un huerto de hongos bien cuidado con un cultivo de laboratorio de los mismos hongos —escribe nuestro amigo Moselio «Elio» Schaechter en su encantador libro titulado En compañía de las setas (1997) —.
Figura 7.2 Pterotermes occidentis (parecida a Heterotermes tenuis).
Figura 7.3 Pterotermes occidentis, simbiontes intestinales de una termita comedora de madera.
Milenios, en realidad millones de años de práctica y selección evolutiva, han desembocado en enormes colonias de termitas que viven en montículos denominados termiteros. Las ciudades de la Macrotermes natalensis, una constructora de rascacielos de Sudáfrica, albergan típicamente de entre uno a dos millones de individuos en cada una de sus grandes estructuras cónicas dotadas de aire acondicionado, que se elevan a más de tres metros por encima del nivel del suelo. El propio nido, construido sobre un espacioso sótano, está apuntalado con columnas. De aproximadamente dos metros de ancho, está construido de tal modo que unas dos terceras partes quedan por debajo del nivel del suelo. El «nido» o recinto donde residen las termitas y sus prósperos huertos de hongos, se encuentra próximo al nivel del suelo y ocupa menos de un tercio del volumen total del termitero. El nido está conectado con el exterior mediante túneles radiales, conductos verticales y por último, por una chimenea central cerrada por su extremo superior. A través de la ciudad de la Macrotermes fluyen unos nueve litros y medio de oxígeno cada hora, de los cuales tan sólo uno y medio son atribuibles a las termitas. Los ocho litros por hora restantes corresponden a la respiración de los hongos en los huertos interiores. El termitero como un todo genera cincuenta y cinco vatios de calor, de los cuales aproximadamente cuarenta y siete son atribuibles al metabolismo de los hongos y tan sólo ocho a las transformaciones químicas en los cuerpos de las propias termitas.
Las termitas suprimen la sexualidad de los hongos, al mismo tiempo que estimulan la producción de «brotes» jóvenes comestibles. Mientras estas macrotermitas se ocupan de podar y cuidar sus huertos de hongos, la formación de setas queda completamente inhibida. Las setas son manifestaciones externas de la vida sexual secreta de los hongos. Tan sólo cuando las termitas agricultoras mueren llegan a brotar setas de los termiteros. Las consecuencias del abandono de sus responsabilidades agrícolas por parte de las termitas son profundas para los humanos. En los termiteros abandonados aparecen exquisiteces tales como la Termitomyces titanicus, una seta de más de un metro de diámetro. Schaechter incluye a las setas termiteras recolectadas en las «ciudades fantasma» de las termitas «entre las más preciadas de los mercados de Nigeria, Zambia y países tropicales de Asia del Sur». De hecho, es tan sólo debido a que las termitas abandonen eventualmente el cuidado de sus huertos como hemos llegado a saber que el hongo Termitomyces, del que aparentemente cabe distinguir numerosas clases, pertenece a los basidiomicetes u hongos productores de setas. Los brotes hinchados y asexuados, el crecimiento en forma de estafilos, están inducidos por la actividad colonial frenética de las termitas. Esta forma túrgida, de extremo hinchado, es única y distinta de los patrones corrientes del crecimiento fúngico. Si esta asociación agrícola entre hongos y termitas fuera realmente permanente y las termitas no abandonaran su residencia, nos veríamos obligados a clasificar a los hongos como «imperfecti». Por razones que no comprendemos, miles de especies de « fungi imperfecti» carecen por completo de fases sexuales. Cultivando esporas antes de la madurez sexual, el experto en hongos Paul Stamets y sus compañeros de Washington State descubrieron un medio que podría liberar a los hogares de termitas y hormigas carpinteras. (Lo ha patentado, ver en la red). Los insectos coleccionan esporas pero, cuando éstas maduran, el efecto fúngico actúa a modo de insecticida natural letal para toda la población. Aquí vemos la razón, al menos de forma superficial, de la comunidad de termitas más activa en agricultura: estas termitas suprimen por completo la actividad sexual de los hongos. Mantienen a los hongos en un constante estado asexual, produciendo alimentos. Resulta interesante observar que también los agricultores humanos suprimen la sexualidad en algunas especies de plantas. Las ramas se injertan antes de florecer, o se seleccionan frutos sin semillas.
Las termitas obreras discurren por los corredores y las grietas de la ciudad fúngica y sus notables propiedades, y salen al exterior para cosechar madera y otras formas de vegetación. Utilizando sus propios intestinos como dilatadas bolsas de transporte, penetran con su cargamento en el termitero por sus múltiples pasadizos y canales. Cuando dan con el huerto adecuado, regurgitan la pasta de madera y hongos, parcialmente digerida. Al principio, naturalmente, esta masa en bruto contiene toda clase de hongos en putrefacción, junto con otros microbios intestinales, pero, de algún modo, el trasero de la termita se convierte en una especie de trasplantador vivo inocuo, o cuanto menos aséptico. La obrera arroja la pasta a medio digerir sobre el suelo de la cámara interior del nido. «Una vez excretado, el micelio fúngico es moldeado en pequeñas esferas, del tamaño de la cabeza de un alfiler pequeño —escribe Schaechter—, Estas esferas, abarrotadas de esporas fúngicas, constituyen la característica más prominente de los huertos de hongos. Para las termitas, la escena debe causar la misma impresión que para nosotros un campo repleto de bejines gigantescos». Esas esferas en forma de cabeza de alfiler constituyen el abundante alimento fúngico que mantiene viva a una ciudad de dos millones de obreros y a sus gobernantes. La reina sedentaria, los soldados de mandíbulas imponentes y las termitas jóvenes recién salidas del huevo deben ser alimentadas «proctodialmente» o «estomatodialmente» (es decir, por medio de contacto de ano a ano o de boca a boca) por las obreras. Las esferas fúngicas son ávidamente devoradas como desayuno, almuerzo y cena.
Los termiteros son ciudades dinámicas y elevadas, construidas y mantenidas por cientos de miles, a menudo millones, de insectos en comunicación constante unos con otros. Cámaras de nacimiento, incubadoras, el equivalente de escuelas, hospitales, salas de luna de miel, talleres y depósitos de cadáveres, todo ello debidamente abastecido de flujos gaseosos y líquidos, actúan a modo de pólizas de seguro con clima controlado para los individuos que sustentan. Al ser atacados por un cerdo hormiguero hambriento o batidos por una torrencial tormenta veraniega, los ciudadanos de estas aglomeraciones de macrotermitas sudafricanas responden con rapidez para restaurar el estado de sus conjuntos residenciales de lujo. El oxígeno circula vivamente por su aire saturado de vapor de agua. El nivel de dióxido de carbono se mantiene entre un dos y un cinco por ciento. Este valor de CO2 es muy superior al 0,032 por ciento de media habitual en la atmósfera terrestre. En la estación seca, la humedad de los huertos de hongos es mantenida gracias a la humedad general del interior del termitero.
Cuando estas fortalezas amuralladas y rematadas por chimeneas se ven amenazadas por un potencial desastre, sus ciudadanos responden como es de esperar: suenan las campanas, aúllan las sirenas y los equipos de especialistas se movilizan. Se pone en marcha una estrategia de realimentación positiva llamada «estigmergia» (de stigma = señal y ergy = energía, literalmente «movido por la señal»). La termita que se encuentra ante el peligro responde de modo reflejo, coge un grano de arena y lo pega en el lugar con una secreción bucal pegajosa.
A esta acción se le denomina «embarrar». Luego libera una alarma química en forma de feromona. Esta sustancia recorre rápidamente los canales y los conductos de toda la ciudad. Las termitas situadas en el lugar del peligro golpean rápidamente las paredes del termitero con sus cabezas. De este modo, la alarma producida en el lugar del problema se difunde rápidamente por la comunidad, desde los pocos individuos afectados hasta las multitudes. Cuando otras termitas detectan el residuo del embarrado de su compañera, embarran también como respuesta. El embarrado simpático se multiplica. La señal feromónica y el golpeteo con las cabezas viaja desde el origen del problema a través de los grandes pilares, de las salas, de las cámaras y de los huertos de la gigantesca morada. Es la estigmergia en acción. La repuesta del grupo es inequívoca: todos sus miembros acuden al lugar del problema. En pocas horas está en marcha la reconstrucción y el boquete queda sellado. Equipos de termitas obreras construyen nuevos canales, nuevas galerías, nuevas columnas y nuevas salas. En palabras de J. Scott Tumer en su fascinante libro The Extended Organism (2000), toda su arquitectura es una «estructura adaptativa». Tumer denomina «homeostasis social» al comportamiento que subyace en el dinamismo de la Ciudad de las Termitas. Describe como sigue la vuelta a la normalidad tras la conmoción: «Una perturbación en la atmósfera de la colonia desencadena una respuesta que la reintegra a su estado anterior a esa perturbación». La tendencia natural de bacterias, protistas, hongos y termitas a unirse para producir estructuras emergentes es igual que la nuestra, sólo que la suya lleva 80 millones de años actuando sobre centenares de especies, mientras que nuestras pretenciosas uniones comenzaron hace tan sólo 8.000 años. Esas fusiones y uniones que creemos exclusivas del género humano tienen, en realidad, numerosos precedentes en el mundo no humano. Las termitas agricultoras no son más que un ejemplo entre otros muchos.
§. Los orígenes independientes de la agricultura
En varias ocasiones a lo largo de la historia del planeta Tierra, los animales sociales han domesticado sus fuentes de alimento. La agricultura se desarrolló entre las termitas, las hormigas y los seres humanos. «Domesticar» significa «llevarse a casa». Algunas asociaciones agrícolas se hicieron aún más íntimas. En ellas las fuentes de alimento no son ya cultivadas muy cerca de casa, sino en el propio cuerpo del consumidor. Narraremos aquí una historia sobre el origen del huerto familiar, y luego describiremos brevemente la relación aún más íntima de la «agricultura corporal». La primera asociación, que ya hemos comentado, entre termitas y hongos sitúa la escena, mientras que la segunda, entre bacterias y protistas de las termitas, atestigua la adquisición de genomas externos. Bacterias alimenticias, algas alimenticias, e incluso ciliados almacenadores de almidón, son criados, respectivamente, en el interior de los cuerpos de algunos protistas, de lombrices verdes y de vacas.
A la lujuriante selva tropical que rodea a la Estación de Biodiversidad del Tiputini sólo se puede llegar bajando por el río desde Coca, tras un viaje de seis horas de duración en canoa, una embarcación fluvial a motor sin ninguna protección. Las termitas, incluyendo la frenética especie subterránea Heterotermes tenuis, abundan en la región, particularmente entre los árboles caídos. Como todos los miembros de la familia Rhinotermitidiae, estas virtuosas de la digestión de la celulosa albergan una dotación muy activa de microbios en sus abultados abdómenes. En su intestino nos encontramos con madera en estado de degradación. A través de los cuerpos traslúcidos de células hipermastigotes, tales como la Trichonympha y algunos de los pirsonínfidos, pueden distinguirse directamente trozos de madera.
Unos centenares de miembros de la atareada colonia de termitas fueron llevados al laboratorio pero sin retirarles de su hogar, un tronco en descomposición. Dos semanas más tarde, de la madera en descomposición emergía lo que parecía ser centenares de gotas de agua perdurables, de medio milímetro de diámetro… sólo que no era agua. La observación mediante microscopio demostró que aquellas «gotas de agua» eran en realidad esporodoquias, término que tuvimos que buscar en el diccionario micológico. Se trataba de paquetes de tejido de un oscuro «fungus imperfectus» de presencia limitada al hemisferio sur. Con la ayuda de algunos amigos conseguimos localizar la escasísima bibliografía existente acerca del misterioso hongo. La presencia de las esporodoquias nos hizo sospechar alguna clase de cultivo agrícola fúngico, tan bien documentado en el caso de las termitas «superiores». ¿Acaso se estaba iniciando el cultivo de hongos entre las termitas comedoras de madera de los trópicos de Sudamérica? ¿Nos encontrábamos ante un nuevo origen independiente de la agricultura?
Andrew Wier, un estudiante graduado, microscopista electrónico y buceador, regresó de pasar dos semanas en la Estación de Biodiversidad del Tiputini durante el invierno del primer año del nuevo milenio. Volvió al laboratorio Margulis extasiado con las maravillas de la selva tropical ecuatoriana. Creía haber reconocido entre los grandes árboles con «contrafuertes» en el tronco algunos representantes de la especie Iriartea, de la familia de las aracáceas. De las ramas y los troncos de muchos Iriartea brotan epifíticamente numerosas bromelias. Algunas cianobacterias de la corteza tiñen de verde su tronco, mientras que hongos blanquecinos les dan la apariencia de pájaros de papel. La diversidad de las palmas lujuriantes es maravillosa. A Wier no le sorprendió demasiado comprobar que el Tiputini estaba a una altura demasiado baja como para poder albergar a la Guanera manicata, una enorme planta simbiótica que encontraremos de nuevo en el capítulo 11. Quedó maravillado con la Avenida Arbórea Amazónica de Bart Bouricious, a cuarenta metros por encima del suelo. Bouricious se gana la vida en un campo extremadamente libre de competencia: construye paseos arbóreos en la cúpula amazónica. El acceso de Bouricious a las copas de los árboles ha sido exhibido incluso en las zonas templadas de Hampshire y en las universidades Williams de Massachusetts. Subir hasta el final de la escalera y observar a los macacos y a los monos araña desde el corredor horizontal, entre la lujuriante vegetación de la selva tropical, constituye una experiencia inolvidable.
En su condición de avezado científico de campo, Wier identificó a los actores —al menos en líneas generales— del escenario ecológico del Tiputini. El reconocimiento de los taxones superiores —palmas, hierbas, cigarras, filodendros, bejucos, líquenes, orquídeas, higos, coníferas, helechos, gusanos poliquetos, crustáceos isópodos, ranas arbóreas, escarabajos, nematodos, peces hueso— no es, por supuesto, demasiado complicado. Sin embargo, la categorización general dista mucho de la identificación precisa de la especie, indispensable para la investigación. Todo cuanto Wier necesitaba para comenzar era el conocimiento general suficiente como para identificar troncos húmedos repletos de termitas comedoras de madera. Para cuando visitó Ecuador estaba estudiando un organismo denominado Staurojoenina, un protista de exuberante complejidad que evita el oxígeno.
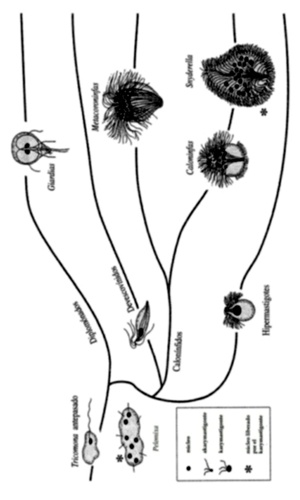
Figura 7.4 Arqueoprotistas: arqueoamebas, diplomónadas, parabasálídas (devescovínidos, Calonínfidos) e hipermastigotes.
El mundo de las termitas, los hongos y los manglares se había ido convirtiendo en el de Wier a lo largo de sus cuatro viajes a Puerto Rico. Justo antes de partir de Ecuador decidió que «su» nuevo protista Staurojoenina pertenecía a la misma especie de otro bien establecido en la bibliografía sobre termitas, llamado Staurojoenina assimilis (figura 7.5) utilizando un microscopio electrónico escáner, Wier descubrió que el protista estaba recubierto por un gran manto de bacterias en forma de bastoncito, una multitud de abigarrados epibiontes. Estas bacterias se extendían por toda la superficie anterior de la ninfa. Wier dedujo que se trataba probablemente de la misma bacteria que la revelada en los espectaculares micrográficos por transmisión de electrones de Staurojoenina, realizados por David Chase a principios de los 80. Wier envió en un vial con alcohol unas cuantas de las termitas soldado muertas de Werle a Rudi Scheffrahn, del Centro de Investigación y Educación de la Universidad de Miami, para su identificación. Scheffrahn es un fenómeno en el reconocimiento de termitas «inferiores» (comedoras de madera), aun cuando se estime que existan una 400 especies vivas e incluso más extinguidas. En su laboratorio, Scheffrahn llegó a la conclusión de que se encontraba ante ejemplares de Heterotermes tenuis, una termita inferior común en los trópicos del nuevo mundo.
Al cabo de una semana aparecieron las pequeñas gotas translúcidas en disposiciones irregulares, alrededor de la madera podrida en las que vivían las termitas. A las dos o tres semanas las gotas alcanzaron el tamaño de cabezas de alfiler, y así permanecieron durante meses. A esta colonia de H. tenuis no parecía importarle la «lluvia» (en este caso de agua destilada), a diferencia de las otras termitas inferiores que conocíamos. El exceso de agua produce inevitablemente que las termitas sucumban a la invasión por hongos y mueran.
Cuando examinamos las gomosas «gotitas de lluvia», nos percatamos al punto de que eran lo más parecido que habíamos visto nunca en la Naturaleza a un cultivo de esporas fúngicas.
Figura 7.5 Staurojoenina: hipermastigote individual compuesto.
La evidencia se acrecentó cuando uno de nosotros (Lynn Margulis) tuvo la fortuna de poder visitar la Estación de Biodiversidad del Tiputini a principios de junio del 2001, invitada por la Universidad de San Francisco de Quito, Ecuador. Con la ayuda del guía nativo Mayer Rodríguez y del profesor de microbiología Gabriel Trueba, pudo visitar a las Heterotermes en hábitat natural de madera empapada. La madera de la palmera Asterocaryum chambina, un majestuoso gigante de entre treinta y cinco y cuarenta metros de altura en su pleno desarrollo, proporcionaba, una vez desplomada, los materiales necesarios para la alimentación y el hogar de las colonias húmedas de las termitas subterráneas comedoras de madera. Allí estaban ellas, gozosamente ocupadas en masticar trozos de palmera, cuando sacrificamos a algunas obreras para indagar en sus secretos. La presencia de una triconínfida, una espirotriconínfida y algunos protistas pirsonínfidos bastó para confirmar que las Heterotermes tenuis de Wier o bien se habían desarrollado entre madera de palmera, o bien lo podían haber hecho perfectamente. Se trataba claramente de termitas «inferiores» comedoras de madera, que vivían en las máximas condiciones de humedad que jamás habíamos visto soportar a ninguna otra termita. Los naturales del país suelen decir que la región del río Napo y de su afluente el Tiputini tiene dos estaciones: la lluviosa y la lluviosa.
Pirozinsky argumentó que, en África, el hongo Deloria palmicola crecía probablemente en la madera húmeda de palmera, particularmente en descomposición. También allí era probable que el Deloria sirviera de alimento a las termitas, aunque él mismo no lo hubiera podido presenciar. Todas las observaciones iban encajando. Un nuevo descubrimiento nuestro vino a sumarse a la evidencia: la abundancia de esporas de Deloria en los intestinos de nuestras Heterotermes. Junto con ninfas digestoras de madera y miríadas de bacterias, sus entrañas estaban repletas de las inconfundibles esporas de tres celdas del hongo Deloria palmicola.
Nuestra dilatada experiencia nos ha demostrado que el cuidado de la humedad es crucial para las colonias de termitas en el laboratorio. Demasiada agua estimula el crecimiento de hongos en los isópteros cautivos. Un pequeño error de un estudiante demasiado voluntarioso que humedezca demasiado la colonia o que no se percate de algún poso de agua, puede significar la muerte de toda la colonia en el fin de semana. Por ello nos sorprendió sobremanera que aquellas colonias de Heterotermes tenuis inundadas de «lluvia» continuaran medrando. Al parecer eran capaces de responder con una estrategia de recuperación al estilo de «el negocio como de costumbre» que, ante nuestra mirada inexperta, se nos antojaba comparable a la descripción de «estigmergia» de J. Scott Tumer. Las agitadas termitas movían sus antenas y, por lo visto, durante el húmedo fin de semana se dedicaban a parchear las partes hundidas de su residencia, aún húmeda. Las inundaciones y las lluvias torrenciales —inferimos— debían ser cosas corrientes en su hábitat ecuatoriano.
La prototaxis es de aplicación a la totalidad de las complejas relaciones entre hongos, termitas, protistas y madera. Estas relaciones comienzan de forma casual, luego se vuelven íntimas y, finalmente, evolucionan a individuos nuevos y únicos.
Tal vez el proverbio de que «la mejor defensa es el ataque» haya sido el primer paso de la estrategia de estas termitas. Los hongos y la lluvia habrían constituido sin duda amenazas constantes para los antepasados de la Heterotermes tenuis de la selva tropical. Sin embargo, aquellas colonias que supieron manejarse entre la constante humedad y comenzaron a comerse a los hongos invasores, lograron sobrevivir y prosperar. Presumiblemente, el hongo Deloria lleva consigo al menos algunas de las enzimas de celulasa necesarias para la descomposición de la lignina. Cuando una Heterotermes tenuis come Deloria se hace con dos por el precio de uno: toma prestados del hongo vivo el nitrógeno consumible, procedente de la quitina, y la proteína de la pared celular del hongo, junto con sus enzimas de celulasa.
Es igualmente probable que las termitas de la selva del Tiputini estén o no hoy en día en camino de convertirse en termitas «superiores». Cuando las examinamos, todas ellas estaban aún repletas de ninfas digestoras de madera. Pero quizás entre estas ecuatorianas se encuentre algún nuevo agricultor de hongos, vacío de protistas celulósicos, listo para dar inicio a la arquitectura urbana. O tal vez no. Estas termitas amantes del agua parecen extremadamente similares a sus antepasados del Mesozoico, que eligieron el cultivo externo de hongos comestibles, al cultivo interno de microbios digestores de madera. Es probable que, durante un tiempo, esos antepasados se alimentaran de las dos formas. Este posible itinerario, desde lo inferior a lo superior, implica tanto a la especiación como a la simbiogénesis. Antes de que las termitas superiores se dedicaran a cultivar hongos en gran escala, en termiteros de medioambiente controlado, algunas termitas inferiores, infestadas de protistas, se defendieron del asalto fúngico domesticando e ingiriendo a sus atacantes. Aquellas termitas aprendieron el truco del crecimiento de hongos en «cultivo puro», promoviendo la formación de esporodoquias gutiformes en cantidades masivas sobre la superficie de sus nidos. Podaron, plantaron y comieron esporas procedentes de esos huertos, exactamente del mismo modo que nuestros antepasados cazadores-recolectores dieran los primeros pasos hacia la agricultura cuidando de (y defecando cerca de, es decir, fertilizando) manchones naturales de vegetales comestibles. Inferimos que las técnicas de cultivo fúngico de las termitas tuvieron que preceder a sus construcciones elevadas. Ese cultivo tuvo lugar primero sobre troncos caídos, en el interior de las umbrías selvas tropicales y en terrenos húmedos, antes de que ese linaje de termitas comenzara a construir sus ciudades criaderos de hongos.
La dependencia de las termitas de la agricultura fúngica superó el punto de no retorno cuando esos insectos perdieron sus simbiontes protistas digestores de madera. Las celulasas —las enzimas degradadoras de la madera— son raras o inexistentes en las bacterias de las termitas.

Figura 7.6 La ciudad de las termitas.
En los estudios sobre las termitas —un mundo dentro de otro mundo— no aparece ninguna bacteria digestora de madera. La solución a la fuerte presión selectiva de conseguir extraer alimento del entorno inmediato, particularmente a partir de madera sólida y resistente, condujo a las termitas a la agricultura. Un grupo de ellas abandonó la madera. Aunque ésta abundaba, la nutrición a partir de ella era demasiado inaccesible. Otro grupo optó por convertir a sus intestinos en una guardería de protistas nadadores, cada uno de ellos recubierto o relleno por genomas bacterianos perfectamente distinguibles, bacterias sea en forma de bastoncitos, de filamentos o de esferas (o los tres tipos juntos), completas con sus genomas intactos. En el caso de estos protistas, que nunca caen en la tentación del sexo, fue la adquisición e integración de simbiontes bacterianos permanentes lo que condujo a la aparición de nuevas especies.
Parte III
El legado planetario
En los tiempos de la caza, que en muchos lugares duraron desde los albores de la Humanidad hasta todo el siglo XIX, eran principalmente los machos humanos quienes corrían durante días en busca de presas que abatir. La pérdida efectiva de calor por medio del sudor se vio favorecida por la pérdida de pelo corporal. Los hombres cazaban en grupos comunicados, mientras que las mujeres se ocupaban principalmente de cosechar plantas y frutos y de cuidar de la progenie. Tal vez algunas de entre ellas (selectas «farmacéuticas ancestrales») se hicieron expertas en el reconocimiento de plantas con propiedades medicinales. En determinados momentos y lugares, algunos machos, frecuentemente en linajes de descendencia directa, fueron preparados para actuar como «padres» de la tribu. Vista retrospectivamente, podríamos decir que esta figura se correspondería con un político-sacerdote-chamán, todo en uno. En nuestros días proliferan los roles sociales, a medida que crece el número de especialidades. Tenemos carniceros, panaderos, candeleros, jugadores de béisbol, mecánicos del automóvil, agentes de seguros y reparadores de televisión. Nuestras escuelas y nuestras universidades registran una creciente diversidad de «campos» académicos. Las ciencias biológicas, por ejemplo, se dividen en un número enorme de grandes subespecialidades: anatomía y ultra estructura, microbiología, biología celular, paleontología, fisiología, genética, biología molecular, etc. Sin embargo, todas ellas estudian evidentemente aspectos de un mismo planeta vivo. Al igual que sucede en la sociedad moderna con la emergencia de un número creciente de oficios, la territorialidad académica refleja el crecimiento inexorable de las poblaciones humanas, junto con la creciente eficacia derivada de la división del trabajo. La especialización forma parte de la respuesta a la pregunta sobre cómo acabó la filosofía convirtiéndose en la ciencia moderna: unos teorizan y otros observan, y las áreas de observación se hacen más pequeñas con cada nueva década, a medida que crece el número de buscadores de datos.
Esa tendencia hacia una creciente especialización no es exclusiva de los humanos. La especialización también aumenta en los ecosistemas, a medida que éstos van desarrollándose.
Cuando un ecosistema boscoso o ribereño crece de nuevo tras un incendio, una inundación o cualquier otra perturbación, su recuperación comienza con poblaciones de crecimiento rápido de organismos del mismo tipo. Todos estos organismos pioneros (determinadas bacterias, algas o hierbas, según el ecosistema de que se trate) se comportan de forma muy parecida: sus poblaciones crecen rápidamente para ocupar cuanto antes el espacio-nicho disponible. Pero, inevitablemente, acaban por enfrentarse a limitaciones del entorno que restringen su crecimiento ilimitado. La falta de espacio, de agua, de fósforo o de otros nutrientes impide que avance el crecimiento de su población. Llegado ese momento, otras especies de crecimiento más lento, que no hubieran podido establecerse en las condiciones iniciales del entorno, se unen a las de crecimiento rápido para hacer un uso más eficiente de los nutrientes, de las fuentes de energía, del agua o de cualquier otra variable del entorno disponible con limitaciones. Estas comunidades de organismos de crecimiento más lento y más estables en un lugar concreto, denominadas ecosistema clímax, acaban por substituir a las pioneras, a veces tras una larga sucesión de etapas intermedias. Estos grupos clímax de plantas, animales y microbios tienden a presentar la máxima diversidad de especies, las interacciones más complejas y los máximos niveles de eficiencia energética en comparación con las comunidades precedentes.
La especialización, sea ésta de ecosistemas, de investigadores científicos o de oficios urbanos modernos, genera organización dentro de poblaciones de grandes números de individuos. Consideremos un ejemplo próximo a nuestro hogar. Una de las poblaciones humanas más densas sobre el planeta se encuentra justamente en nuestra pequeña y bucólica ciudad de Amherst, en Massachusetts. La densidad de la población estudiantil en el área residencial del sudeste de la Universidad de Massachusetts es de 5.400 personas por cuarto de milla cuadrada [13]. La ciudad de Nueva York tiene «tan sólo» 1.500 residentes por la misma unidad de superficie. La razón es simple: las ciudades como Nueva York están densamente pobladas por su configuración en rascacielos de oficinas y bloques de apartamentos, gran parte de todo lo cual se queda vacío por la noche. Los dormitorios de la ciudad estudiantil en cambio, el más alto de los cuales no supera los veintidós pisos, soportan un enorme número de estudiantes porque éstos viven apretados. Las habitaciones de dos camas carecen de cocina, de baño privado y de espacio abierto. Los neoyorquinos, en cambio, gozan de varios parques —particularmente del Central Park— así como, en general, de más de medio dormitorio per capita. En los enormes y elevados bloques de dormitorios en forma de U, edificios especializados están dedicados a proporcionar los servicios de comedor a centenares de estudiantes. El conjunto como un todo no dispone de espacio abierto, de campos de deporte ni de salas comunes. En cambio, los alojamientos de Manhattan suelen tener baño, cocina y áreas comunes en cada unidad, y no hay edificios especializados en servicios.
La especialización tiene efectos contrapuestos. Aunque indudablemente eficaz para la organización de grandes cantidades de individuos, la especialización nos aleja del sentido de la totalidad. Puede que la ciencia, en cuanto actividad internacional, sea hoy más eficaz que en el pasado, pero los académicos multidisciplinares al estilo de los del Renacimiento, familiarizados con la práctica totalidad del conocimiento, son cada vez más escasos, cuando no completamente inexistentes. Nadie puede ya estar familiarizado con todas las ramificaciones de la cultura. Sin embargo, tal vez sea posible contemplar el todo. Quizás sea ésta una de las razones del crecimiento de la ciencia popular, que le permite al ciudadano corriente una visión general, al menos desde el punto de vista de las ciencias naturales, y obliga al científico o al escritor científico a describir adecuadamente ése todo. Probablemente ésta sea también la razón del atractivo de la ciencia de la evolución: la evolutiva es una ciencia de conexión.
Antes de que esta ciencia comenzara a dar sus primeros pasos en la comprensión humana, la noción funcional de la totalidad era una cuestión más bien reservada a las religiones: el dios único de las religiones monoteístas, tales como la cristiana, la judía o la islámica. Se daba por sentado que cada forma de vida, cada especie nombrada, había sido creada individualmente por Dios, reflejando en sí misma una variación más del plan divino. El intérprete humano de ese diseño, fuera éste sacerdote o santo, ocupaba un lugar de observación privilegiado. Pero Lamarck y Darwin plantearon una totalidad más realista. Sentaron la base para la comprensión del vínculo entre todas las especies y todos los organismos, no ya a través de una deidad omnipoiética, sino simplemente a través del tiempo. El genio de Darwin, así como de tantos otros historiadores naturales, criadores y geólogos, cuyo trabajo tanto él como sus sucesores sistematizaron, consistió en demostrar que todos los organismos hoy vivos están conectados, a través del tiempo, por antepasados comunes.
Las personas más cultas creen comprender las plenas implicaciones de esta visión conectiva. Nos permitimos sugerir que algunas de las consecuencias del concepto darwiniano de evolución han pasado desapercibidas. Vemos una relación entre el crecimiento de un animal a partir de un óvulo fecundado y el desarrollo de la biosfera a partir de las primeras células. Ambas entidades se desarrollan y evolucionan por medio del cambio a través del tiempo. Tanto el niño que crece como la biosfera que evoluciona se parecen en que son la expansión de distintos ecosistemas. El cambio de los seres vivos a través del tiempo requiere el incremento del número de células, acompañado de la muerte celular. Aunque numerosas personas cultas parezcan sentirse a gusto con la idea de que evolucionamos a partir de especies parecidas a los simios actuales, la idea de que, en última instancia, somos descendientes de bacterias —es decir, gérmenes— no parece ser tan popular. Que nuestro propio cuerpo esté masivamente poblado por abigarradas aglomeraciones de células, clones naturales de células nucleadas que nacen y mueren sin cesar, es una realidad que pocas veces se señala. Pero, a pesar de ello, provenimos de microbios simbióticos, de células nucleadas especializadas, de animales en forma de gusanos y de peces dipnoos, de reptiles y de insectívoros, de protosimios y de monos antropoides, en suma, de una forma de vida tras otra. Desde que ocurriera la primera manifestación de vida sobre el planeta, en ningún momento se ha interrumpido nuestro linaje ni se ha detenido el sistema.
§. El espacio de Vernadsky, el tiempo de Darwin
Establecer la relación entre nosotros mismos y el panorama general de la historia del planeta requiere una visión que se extienda más allá de nuestros antepasados simiescos, e incluso de los mamíferos y los reptiles que les precedieron. Los microbios fueron la primera forma de vida, los cambios en sus comunidades constituyeron la base de la primera evolución. La transformación de multitudes de células en organismos individuales mayores, aunque omitida de la teoría evolutiva zoocéntrica clásica, forma parte del legado darwiniano, que no es otro que la conexión de todo lo vivo a través del tiempo.
El legado de Vernadsky consiste en la conexión de todo lo vivo a través del espacio.
Vladimir I. Vernadsky (1863-1945) fue un individuo notable, cuya contribución a las ciencias —incluyendo la extensión de la biología a otras ciencias— es mucho más apreciada en el Este que en el Oeste. En la desaparecida Unión Soviética existen unas catorce instituciones que llevan su nombre. Su rostro barbudo puede verse en sobres aéreos y sellos de correos. Tal vez el ateísmo de la Unión Soviética facilitó el clima científico e intelectual adecuado para que las ideas de Vernadsky fueran fácilmente asimilables. En su visión de que la vida constituye con mucho la fuerza geológica más importante, así como de que el planeta Tierra es una entidad única sustentadora de vida, Vernadsky iba muchos años por delante de su tiempo. A pesar de que fuera el geólogo suizo Edouard Suess quien acuñara el término «biosfera» (literalmente, esfera de vida) para complementar a la «hidrosfera» (parte acuática de la Tierra) y la «atmósfera», fue Vernadsky, con su obra La biosfera (1926 [14]), quien llevó el término a su uso generalizado. Esta obra fundamental fue traducida al francés en 1929, pero su versión completa no estuvo disponible en inglés hasta 1998. Vernadsky, químico especializado en cristalografía, mantuvo un interés constante por la estructura de los minerales. Era sin duda un científico monista con intuiciones novedosas. A diferencia de los vitalistas, que mantenían que tenía que haber alguna propiedad especial, exclusiva de la vida, que le confiriera capacidad para pensar y actuar por sí misma, Vernadsky contemplaba la vida como un producto natural de un universo químicamente activo. Los primeros vitalistas argumentaban que la vida tenía que estar constituida por una sustancia particular, que la distinguiera de la materia no viva. Sin embargo, después de que en 1828 Wohler consiguiera sintetizar en su laboratorio la urea, relacionada químicamente con el ácido úrico de la orina —compuesto químico que contiene nitrógeno, oxígeno y carbono—, el argumento de que la vida estuviera constituida por una sustancia especial ya no podía defenderse. Vernadsky llevó el concepto un paso más allá: contempló la vida como un fenómeno global enteramente natural. De hecho, la propia palabra «vida» contiene una connotación vitalista, la de una entidad particular, que no puede ser reducida ni a la química ni a la física. Para evitar esta connotación Vernadsky utilizó decididamente la expresión «materia viva» en lugar de «vida».
Influido por el trajín de armas, hombres, carros de combate y aeroplanos a escala global, con motivo de la primera guerra mundial, Vernadsky veía la vida (incluyendo la humana) como un mineral complejo. La materia viva era para él una especie de «agua animada». A modo de un fluido multicompuesto casi sólido, la vida era una forma de materia altamente cargada de potencial, rica en energía y creativa. Con impurezas minerales y en una fase concreta, la vida es en su mayor parte, sin duda alguna, agua salada. A lo largo del tiempo evolutivo —argumentaba Vernadsky— se fueron implicando cada vez más elementos químicos en la circulación global de materia viva, tendiendo a incrementarse los niveles de circulación de carbono, hidrógeno, nitrógeno, oxígeno, azufre y fósforo. Ese mineral complejo que es la vida, es también transportable (por el viento o el agua, por ejemplo) y auto transportable. Las langostas, por ejemplo, caen sobre los cultivos de grano y los devoran, y luego se elevan de nuevo transportando en horizontal sobre la Tierra su cargamento de agua salada rica en materia orgánica. Esta clase de poblaciones masivas móviles de insectos eran, en la visión de Vernadsky, «montañas voladoras».
Según Vernadsky y Andre Lapo (1987), su paladín moderno, la materia viva constituye la fuerza geológica más poderosa. La parte activa de la biosfera, una fina capa de vida de veinte a treinta kilómetros de espesor, abarca desde las esporas microbianas de la atmósfera a los ecosistemas de los abismos oceánicos. La parte activa de la superficie terrestre se asemeja a la corteza de un árbol en crecimiento. La madera del árbol no está realmente muerta, sino formada por tejido otrora vivo, integrado en el sistema aún activo de transporte de agua y azúcar del corazón del árbol. La parte del paisaje que percibimos como mineral —arena, escombros, mineral de hierro— fue otrora moldeada por las actividades de la vida, o pasó por una fase de producción en el interior de alguna célula. Es el testigo mudo de «biosferas pasadas». Al igual que gran parte del granito de todo el planeta, los acantilados blancos de Dover son microbios formadores de esqueletos fosilizados. El suelo fértil, uno de los primeros objetivos de interés para Vernadsky (que también interesaría a Darwin, que escribió una monografía sobre cómo las lombrices contribuían a su formación), no existiría de no ser por la intensa actividad biológica.
No existe suelo[15]sobre la superficie de Marte o de Venus, sino un escombro denominado «regolita». Húmedos, fértiles y repletos de células protistas y de otros organismos, los suelos son una clase de tejido exclusivo de nuestro planeta vivo. Su composición viene determinada por la circulación global de elementos clave. Habida cuenta de que ningún organismo individual es inmortal, así como de que los materiales de los que estos organismos están compuestos son limitados, los elementos químicos cruciales (carbono, oxígeno, azufre, fósforo y nitrógeno) son constantemente reciclados por todos los ecosistemas, a escala global. Los modelos informáticos de geoquímicos y bioquímicos demuestran la circulación de carbono, oxígeno, agua y otros compuestos químicos sobre la superficie del planeta. La construcción de modelos lo más exactos posible se ha convertido en los años recientes en un objetivo para geoquímicos y bioquímicos, después de que Vernadsky y Lapo centraran el interés sobre la íntima relación entre vida terrestre y geología. La tendencia hacia la vinculación entre biología y geología se ha ido reforzando, particularmente después de la publicación del original trabajo de James Lovelock. Su brillante presentación de la «hipótesis Gaia» fue un llamamiento al despertar de la comunidad científica a la consideración de «la vida y el medio ambiente como un sistema único, estrechamente acoplado», en palabras del propio Lovelock. Completamente independiente del legado de Vernadsky, Lovelock, químico atmosférico, midió discrepancias asombrosas en la atmósfera terrestre, cuyas anomalías vinculó a las actividades de la vida sobre el planeta. Desde finales de los 50, Lovelock ha ido refinando, expandiendo y perfeccionando descripciones de nuestro planeta acuático y único. De forma creciente, los constructores de modelos se han ido percatando de lo que la especialización académica tiende precisamente a oscurecer: la vida —parte integrante del «medio ambiente»— no puede ser ignorada en ninguno de esos modelos. La materia circula alrededor de la biosfera el mismo modo que lo hace por nuestro cuerpo. Tanto la biosfera como el vertebrado contienen partes duras minerales —huesos, dientes, escamas, pelo— que no están, en sí mismas y por sí mismas, vivas.
Esta analogía entre huesos de mamíferos, árboles y el planeta vivo se extiende más allá de lo que pueda parecer a simple vista. Si bien la vida no puede seguramente reclamar influencia alguna sobre el núcleo terrestre en fusión, probablemente juegue un papel prominente en fenómenos aparentemente tan geológicos como la presencia de agua sobre la Tierra —mantenida gracias a que las bacterias, las plantas y las algas se encargan de retirar de la atmósfera los gases de invernadero—, la meteorización —debida a que bacterias y hongos pulverizan y digieren las rocas sujetas a erosión— o incluso la tectónica de placas, como veremos más adelante.
La hipótesis Gaia de James Lovelock ha representado un gran avance desde que Vernadsky reconociera a la vida como fuerza geológica. Lovelock postula que tanto la composición química de la atmósfera, como su temperatura global, la salinidad de sus océanos y la alcalinidad de la superficie de éstos (pH 8,2), no son parámetros aleatorios, sino que presumiblemente vienen regulados por el metabolismo de la suma de la vida sobre la Tierra. Esta clase de modulación global no significa que la superficie de nuestro planeta sea el equivalente a un organismo, porque, a diferencia de la biosfera, no puede sobrevivir de sus propios residuos ni respirar sus propias excreciones gaseosas. Sin embargo, la superficie de la Tierra sí presenta algunos rasgos propios de los organismos. Está construida en gran medida a base de células que se reproducen, toma sus nutrientes del agua y produce incesantemente residuos. Ambos entran en asociaciones ecológicas, en ocasiones simbióticas, absolutamente necesarias para el reciclado de residuos, lo cual determina que el reino celular se expanda. El resultado consiste en que, con el paso del tiempo, el medio ambiente se vuelve cada vez más organizado, diferenciado y especializado.
Partes del medio ambiente pueden acabar finalmente organizadas dentro del cuerpo o como extensión del mismo. No solemos considerar al calcio de nuestros huesos como parte del «medio ambiente», sin embargo, así es exactamente como se originaron los huesos y muchas otras partes duras en la historia de las células nucleadas. El calcio, residuo tóxico sobreabundante en el agua de mar, tenía que ser retirado de algún modo. Metabolizado en forma de carbonato cálcico (CaCO2) o de fosfato de calcio, sustancias que se iban acumulando en las células, acabó siendo usado para la construcción de protecciones y estructuras de refuerzo. Entre las colonias para las que estas partes duras resultaron ser de utilidad, se cuentan nuestros antepasados acuáticos. Entre otros estaba también los antepasados de los moluscos, los artrópodos, un planctónico con cáscara dura conocido con el nombre de diatomea, foraminíferos y cocolitofóridas cuyas cáscaras, precipitándose en una lluvia constante sobre el fondo marino durante millones de años, acabaron por crear los lechos arenosos y cretáceos que, con el paso del tiempo, se convertirían en las rocas blancas de Dover (para más detalles ver el famoso ensayo de T. H. Huxley titulado «Sobre un trozo de yeso»).

Figura 8.1 La Tierra de los astronautas del Apolo, vista desde la Luna.
El proceso de incorporación o reutilización de residuos, de crecimiento y reciclaje, de utilización económica de unos recursos limitados en un espacio cerrado, reúne el concepto de Gaia de que el planeta Tierra tiene una fisiología, con el énfasis de Vernadsky sobre la materia viva como extensión de la geología. Mientras que Vernadsky demuestra la naturaleza mineral de la materia viva, desconstruyendo así la jerarquía entre la vida y la no vida terrenales, Lovelock hace otro tanto sacando a la luz diversos comportamientos fisiológicos de la biosfera como un todo. Los puntos de vista de ambos científicos reflejan la interconexión espacial de los sistemas termodinámicamente abiertos, que denominamos materia viva, con el medio ambiente del que obtienen su energía. La química de nuestra atmósfera no es en absoluto aleatoria. Está directamente relacionada con la respiración —la absorción y expulsión de gases— de muchos trillones de células. Cada célula, cada cuerpo, cada ciudad, intercambian gases con la atmósfera las aguas y los suelos del planeta.
La revolución de los 60 y los 70 demostró, sin lugar a dudas, que la Tierra está cubierta por placas litosféricas enormes, húmedas y móviles. Con anterioridad a la definición de la teoría de placas tectónicas como explicación para la deriva continental, se daba por cierto lo evidente: que bajo nuestros pies la Tierra era sólida, que el suelo estaba donde siempre había estado y que, a excepción de los terremotos, mares y tierras mantenían inmutable su distribución. Compuestas básicamente por basalto, más pesado, y granito, más ligero, sabemos hoy que estas placas inmensas de roca sólida se crean en los bordes en recesión y se destruyen donde sus bordes colisionan o se sumergen violentamente.
La tendencia de las placas continentales a desplazarse lateralmente sobre el manto en fusión, también puede relacionarse con la vida. La presencia de agua líquida sobre la Tierra durante centenares de millones de años ha sido tradicionalmente atribuida a la buena suerte de nuestro planeta. Sin embargo, el agua de la Tierra podría muy bien haberse disipado al espacio hace mucho tiempo, como parece haberles sucedido a nuestros vecinos Marte y Venus. En ausencia de las actividades recicladoras de los seres vivos y de su trabajo en la retirada de dióxido de carbono de la atmósfera, la Tierra también podría haberse quedado sin agua. El metabolismo, el crecimiento y la reproducción de células la retuvo. La fotosíntesis utilizó parte del gas de invernadero dióxido de carbono de la atmósfera para producir toda clase de compuestos orgánicos, manteniendo así al planeta en una temperatura suficientemente fresca como para que su vapor de agua no escapara al espacio. Sin agua que lubricara el movimiento de las placas tectónicas, tal vez éstas se habrían detenido. La superficie húmeda viva del planeta mantuvo, pues, las condiciones medioambientales características de un Sistema Solar primigenio. Tal y como Vernadsky apuntara y Lovelock corrobora, la vida es un fenómeno planetario.
Podemos ya comenzar a ver cómo el espacio de Vernadsky y el tiempo de Darwin están conectados al resto del universo físico. A través del metabolismo y de la reproducción, la vida sobre la Tierra tiende a mantener unos estados medioambientales característicos de las primeras etapas del desarrollo del planeta. Nuestro propio cuerpo orgánico y acuoso, al igual que el de los animales, las plantas y los microbios, es una especie de cápsula del tiempo que contiene el entorno químico de la Tierra tal como era hace tres mil millones de años. El pasado pretérito está preservado gracias a los prodigiosos flujos de energía solar, capturados por la fotosíntesis y por profundas reacciones químicas calientes en las entrañas del planeta. El medio ambiente en el que ocurre la evolución es dinámicamente estable y autorregulador. Mantenida en gran medida por las interacciones químicas y biológicas de los miembros de las comunidades microbianas, la estabilidad fisiológica de la biosfera azulada requiere un flujo incesante de energía procedente del exterior del sistema. La vida está ligada al planeta Tierra y a la célula, pero es también un fenómeno geológico y solar.
§. Las edades de Gaia: arqueozoico, proterozoico y fanerozoico
Ponerle por nombre Gaia a su hipótesis fue otro de los golpes de genio de James Lovelock. El nombre (tomado de la diosa Tierra de la antigua Grecia), sugerido a Lovelock por el novelista William Golding, vecino suyo en Cornualles, resulta sin duda mucho más atractivo que el de «La Tierra como sistema químico homeostático». El planeta Tierra es un sistema fisiológico —argumenta Lovelock— y, como en cualquier sistema fisiológico, los parámetros de su regulación —temperatura, flujo y composición de gases, sal, concentraciones ácidas y alcalinas— pueden ser detectados. Son dinámicos y estables. La fisiología es fácilmente inferible a partir de mediciones. Al darle al planeta el nombre de la diosa griega, Lovelock, en un verdadero acto de bautismo, convirtió a su idea en memorable, tanto para la ciencia como para sus enemigos. Logró personalizar el objeto de estudio y centrar la atención de sus colegas en sus características sobresalientes. En la mente de muchos, «Gaia» quedó reducida a un eslogan: «la Tierra es un organismo gigante», de hecho un organismo «femenino». ¿Cómo —protestábamos ante Lovelock— podría ser la Tierra un «organismo», cuando ningún organismo puede vivir de sus propios residuos? ¿Por qué insistes en promover la imagen de la Tierra como un ser único y gigantesco —le recriminábamos— alejándote de la ciencia y atrayendo hacia a ti la potencial enemistad de respetables colegas? ¿Por qué no decir que Gaia es un ecosistema gigante, que reconoce la pluralidad de los seres que lo componen?, le sugeríamos. Pero él insistía. El término «ecosistema» le parece engorroso, difícil de definir y completamente opaco para quienes no estén familiarizados con la ecología. Es más, se atreve a defender su metáfora de la diosa. «Si consigo que piensen en la Tierra como en un ser vivo —explica— aprenderán a respetarla y a amarla. Si el planeta no es más que un montón de rocas, no hay inconveniente en darle puntapiés y maltratarlo. A nadie le importa. Por supuesto, Gaia no es un organismo único ni ninguna diosa, pero está viva y merece nuestra comprensión y nuestra reverencia». Las edades de Gaia (1988), el mejor libro de Lovelock, explica de forma cautivante el desarrollo de la hipótesis Gaia —ahora desarrollada ya como teoría Gaia—, así como el desarrollo del propio planeta. Desde el nacimiento del Sistema Solar al que pertenece, a través de su adolescencia y hasta su madurez, podemos seguir de la mano de Lovelock el desarrollo de nuestro planeta como sistema vivo. La ciencia y los científicos nos explican cómo fueron desvelados sus secretos más íntimos. Se nos presenta el historial médico de los 4.000 millones de años de edad de la Tierra, el seguimiento de sus numerosos cambios y de sus etapas de maduración, e incluso un atisbo de su probable envejecimiento y muerte. La analogía que Lovelock establece entre nuestras propias secuencias de desarrollo y sus sensibilidades predecibles, y los sistemas reguladores que él denomina la fisiología de Gaia, funciona realmente. Aunque «geofisiología» suena más científico, lo cierto es que «Gaia» ha arraigado. «Gaia» es y «Gaia» seguirá siendo. Gaia es el sistema interactivo de la superficie terrestre, alimentado por la energía solar y por gradientes de energía geotérmica, que mantiene la temperatura alrededor del los 18º C. Frente a las amenazas ácidas, mantiene su alcalinidad en un pH de 8,2. La presión atmosférica de sus gases reactivos (por ejemplo del oxígeno, al 21%) está lejos del equilibrio. La nueva ciencia gaiana, en gran parte bajo el manto de la denominación CST Ciencia del Sistema Tierra)[16] es sólida. Para más detalles ver «Gaia y la colonización de Marte», «Gaia y filosofía» y «Una buena palabra de cuatro letras» en la Parte III de Slanted Truths (1999), donde explicamos con detalle la hipótesis Gaia y sus implicaciones para nuestra vida.
Inicialmente, la teoría Gaia incluía a tres sistemas activamente mantenidos: la temperatura, la composición química de los gases reactivos (en particular del oxígeno) y la acidez y alcalinidad. Los nuevos avances en la misma tratarán de dar respuesta a las preguntas que la Gaia de Lovelock ha planteado, entre las que se cuentan la «Gaia acuática» o « ¿Ha sido el agua retenida sobre la Tierra (a diferencia de Marte y Venus) debido a la presencia de vida sobre ella?», el tamaño de Gaia (« ¿Hasta dónde llegan la vida y sus compuestos orgánicos hacia el interior de la Tierra?») Y « ¿Es la concentración de sal (y de otros minerales) un fenómeno exclusivamente gaiano?». Acuden igualmente a la mente otras preguntas, como si el granito es o no una roca gaiana, o si la distribución en el tiempo y el espacio de las grandes formaciones férricas está o no directamente relacionada con la génesis y el desarrollo de la vida. Como el propio Lovelock ha señalado en más de una ocasión, el poder de una buena teoría científica consiste menos en aquello que explica definitivamente, que en las preguntas, las observaciones oportunas y los experimentos que suscita. Bajo este criterio, Gaia es una buena teoría científica. La personificación de Gaia en el libro de Lovelock sobre sus «Edades» no es una argucia editorial, es una ayuda a la diseminación de los principios básicos de una idea compleja.
La metáfora de Gaia como una antigua diosa de carácter fuerte le permitió a Lovelock escribir sobre la historia de la Tierra de forma accesible al gran público. Tal vez algunos de nuestros lectores no estén familiarizados con la «división entre tiempo y rocas», la gran escala temporal que organiza cantidades inmensas de datos geológicos. En este caso resultará de utilidad la visión gaiana del tiempo remoto. Los «eones» son las divisiones de mayor tamaño dentro de esta escala temporal. Tras el eón Hadeano (infernal) [17], anterior a la existencia de la vida, siguen sólo tres eones: el Arqueozoico, el Proterozoico y el Fanerozoico. Necesitaremos estar familiarizados con ellos para poder comprender los orígenes de las especies. Como sistema vivo, Gaia nació al principio del eón Arqueozoico, hace unos 4.000 millones de años. Este eón fue una época de enorme actividad tectónica y meteórica, en la que la corteza de la Tierra fue solidificándose a partir de rocas en fusión y la liberación de gases del interior del planeta creó su atmósfera. Este eón duró 1.500 millones de años. La mayoría de las rocas del eón Arqueozoico son basálticas y sus organismos son bacterias.
Durante este eón la Tierra tenía poco oxígeno en sus rocas, en su aire y en su agua. Hace unos 2.500 millones de años se produjo un gran incremento en el oxígeno, que marca el inicio del eón Proterozoico. Unos 1.800 millones de años atrás el oxígeno abundaba, las rocas graníticas continentales dominaban la escena y aparecieron las primeras células nucleadas. Desde el punto de vista biológico, el Proterozoico es la época de las bacterias aeróbicas y de los protoctistas. Estos dos primeros eones, los eones del microcosmos, terminaron hace 541 millones de años, en la base del período geológico Cámbrico. El eón Fanerozoico, el más breve de los tres y el más conocido, comienza en la frontera entre el Precámbrico y el Cámbrico. Esta frontera está ahora oficialmente establecida en Mistaken Point, en la península de Avalon, en Terranova, Canadá. Este lugar, en el que los fósiles del Cámbrico (eón Fanerozoico) aparecen sobre una plétora de fósiles del eón Proterozoico (la biota ediacárea), es relativamente accesible. Una localidad fosilífera aún más espectacular, como el río Aldan, en Siberia, resulta inaccesible para quienes carezcan del tiempo, del dinero, de las embarcaciones grandes y de las habilidades técnicas de escalada necesarios. En muchos otros yacimientos rocosos de frontera entre el Precámbrico y el Cámbrico, la conservación es deficiente, faltan las secuencias de sedimentación o el acceso es prácticamente imposible. El eón presente comenzó con la famosa explosión de las partes duras de los animales: placas quitinosas, cáscaras, caparazones de los trilobites, algas cubiertas de carbonato cálcico y pellas fecales. En museos de todo el mundo se presenta erróneamente al Fanerozoico inicial como el inicio de la vida. Entre las divisiones de este eón, bien conocidas, se incluyen el Paleozoico, la era de los animales marinos con cáscara, el Mesozoico o era de los reptiles, y el Cenozoico o era de los mamíferos. Sin embargo, Gaia era ya toda una matrona madura cuando comenzó el Fanerozoico. Unos 450 millones de años atrás había desarrollado su manto verde: bosques y plantas bajas habían comenzado a decorar un paisaje antes dominado exclusivamente por algas y cianobacterias.
Para cuando aparece el eón Fanerozoico, hace unos 541 millones de años, Gaia muestra ya muchas huellas de envejecimiento. Ya ha sufrido la mayor amenaza a su supervivencia desde su tumultuoso nacimiento, en los inicios del eón Arqueozoico. Ese acontecimiento fue muchísimo más grave que la desaparición de los dinosaurios y de gran parte de los pobladores vivos del planeta, tras el impacto del meteorito de Chibchulub de hace 65 millones de años y su subsiguiente cambio climático. Incluso la desaparición del 75 por ciento de los principales taxones, hace 225 millones de años en la llamada frontera P/T —extinción permotriásica—, se queda en nada al compararla con esta otra amenaza. La propia vida de Gaia, el sistema fisiológico planetario entero, se vieron amenazados de muerte cuando la mayor de todas las glaciaciones se enseñoreó del planeta: la «Tierra Bola de Nieve» de hace unos 600 millones de años, de la que existen pruebas de finales del Proterozoico en todo el mundo. Los glaciares llegaron a penetrar incluso en las regiones ecuatoriales. Las rocas de carbonato se amontonaban en enormes estratos y el dióxido de carbono se acumulaba bajo los hielos hasta que, debido a la acumulación de gases de invernadero, fruto a su vez de la escasa fotosíntesis, la atmósfera se fue calentando y los hielos se fueron fundiendo. Este ciclo se repitió varias veces, hasta que Gaia lograra finalmente prevalecer.
Unos cien millones de años más tarde, es decir, hace unos 541 millones de años, la Tierra bullía con formas de vida de mayor tamaño, más diversas y más sorprendentes. La gran ecóloga anglo-estadounidense G. Evelyn Hutchinson describió la historia de Gaia como una obra evolutiva en el teatro de la evolución. El teatro —el planeta de Gaia— sigue ahí, pero los actores del drama ecológico van cambiando. Unos se van por su propio pie, otros son despedidos. Otros nuevos se forman por medio de combinaciones y alianzas entre los antiguos. Mutaciones aleatorias y otros cambios genéticos aguzan y refinan esas nuevas alianzas. La obra ecológica sobre el escenario evolutivo sigue en marcha con jóvenes actores noveles y con veteranos reubicados, cambiando incesantemente sus papeles en escenas escritas por autores y poetas siempre nuevos. Merece la pena tanto observar la acción sobre Gaia como participar en ella. Tal vez siga en escena durante otros 4.000 millones de años, cuando finalmente morirá porque el Sol también habrá muerto. Hasta que caiga ese telón final, la obra seguirá desplegándose y Gaia seguirá prosperando.
Capítulo 9
Eucariosis en un mundo anóxico
El Thiodendron de Dubinina y la sangre de DennisDenominamos «eucariosis» a la transición desde las bacterias a nuestras células eucariotas. En la historia de las células vivas con núcleo, las células eucariotas aparecen por primera vez en los registros fósiles del eón protorozoico. La primera evolución de seres vivos formados por una sola célula nucleada —protistas— dejó descendencia tras de sí: amebas, euglenas y ciliados tales como los euplotas o los paramecios. Todos los organismos visibles —plantas, animales y microbios «grandes»— están compuestos por muchas de estas células. Expresado de forma simple, la eucariosis se refiere a la mayor etapa evolutiva jamás ocurrida en la historia registrada de la vida. Entre bacterias y eucariotas no hay eslabones perdidos, ni en los fósiles ni en la propia vida. La repentina aparición de las eucariotas en la escena de la evolución no fue gradual, sino genuinamente discontinua. ¿Cómo sucedió? ¿Cómo se relaciona el origen de las células nucleadas con nosotros, los humanos?
La quimera de Gupta
La liberación del núcleo
La mayoría de personas suscriben de forma inconsciente cierta clasificación entre seres vivos. La vida en el mundo se divide en tres grandes categorías: animales, plantas y gérmenes. (Algunas personas suponen, al menos tácitamente, cuatro categorías: animales, plantas, gérmenes y personas, aunque no es probable que quienes niegan la naturaleza animal de las personas estén leyendo este libro). Toda persona con cierto nivel de cultura apoya la investigación médica que demuestra que los humanos somos mamíferos como las ratas o los ratones. Cualquiera que eche un vistazo estará de acuerdo con que los humanos tenemos un esqueleto óseo con vestigios de cola, pulgares opuestos, visión tridimensional, dientes y otras características que nos sitúan inequívocamente entre los mamíferos primates. Incluso con conocimientos científicos escasos, el lector reflexivo reconocerá instantáneamente la naturaleza cordada (espina dorsal, sistema nervioso y cerebro), mamífera (pelo, glándulas mamarias) y primate (manos de chimpancé, dientes, postura y conducta social) de los humanos. Por supuesto, todos los humanos pertenecemos hoy a la especie Homo sapiens porque todos nosotros —al menos en principio— podemos aparearnos y producir descendencia potencialmente fértil.
Los microbios (las bacterias, los protoctistas y los hongos más pequeños) fueron descubiertos por Antony van Leeuwenhoek, un tapicero holandés que se construía sus propios microscopios allá por el año 1675. El químico francés Louis Pasteur reveló un mundo microbiano muy distinto del ordinario de los organismos mayores. Las prácticas de Pasteur determinaron la emergencia de una nueva ciencia, donde la relación de bacterias y levaduras con los humanos fue estudiada intensivamente. ¿Tenía algo que ver la presencia de aquellas pequeñas manchas crecientes (poblaciones de bacterias) con la fiebre infantil? ¿Sería acaso un virus el causante de la glosopeda? ¿Estaba tal vez relacionada la aparente capacidad de aquellas burbujitas para producir alcohol, con la tendencia del jugo de uva a convertirse en vino? Incluso hoy en día, no es fácil encontrar en la bibliografía médica, biológica y de nutrición, un conocimiento básico acerca de la vida de los microbios. Los «datos» se enseñan en una base de necesidad-de-conocer. La Spirulina, por ejemplo, es un aditivo nutricional común. Aunque se trata inequívocamente de una bacteria fotosintética productora de oxígeno, lo cierto es que se comercializa bajo el nombre de «alga». La sinceridad no es siempre la mejor de las políticas posibles: promocionar deliberadamente la bebida de una solución bacteriana podría ser una locura comercial.
La similitud entre plantas y animales no es intuitivamente evidente, pero están mucho más relacionados entre sí que cada uno de ellos con las bacterias. Existe en la Naturaleza una discontinuidad mucho mayor entre las bacterias y ambos que entre cualquier planta y cualquier animal. Se trata de un concepto difícil incluso para los naturalistas y otros científicos que conocen la existencia del mundo microbiano, aunque desde finales del siglo XIX, el trabajo de centenares de científicos ha ido convergiendo sobre esta idea relativamente nueva. La mayor discontinuidad sobre el planeta no es la que media entre animales y plantas, sino la que separa a procariotas (bacterias sin núcleo encerrado en membrana celular) y eucariotas (todos los demás seres vivos formados por células nucleadas). La historia detallada de esta enorme discontinuidad está relacionada con los orígenes de las especies.
El relato de la historia de las células es el del modo en que las células comenzaron a generar especies. Para entender por qué, revisemos con mayor detalle una cuestión profundamente relacionada con el origen de las especies: el aumento de descendencia en las bacterias, paralelo al aumento de la aparición de especies. En el caso de organismos que se reproducen por medio de apareamientos fértiles, las especies constituyen entidades reales fácilmente reconocibles. Los chimpancés se aparean con chimpancés, los ovarios de las flores del maíz son fertilizados por el polen de maíz, el esperma de rana fluye sobre los huevos de rana y los convierte en huevos fértiles de los que nacen renacuajos. Únicamente los miembros de una misma especie copulan, fertilizan, fecundan o lo que sea, para producir nuevos miembros de esa misma especie. Pero, antes de las primeras células nucleadas, el planeta albergaba únicamente a bacterias, y con ellas, las reglas aparentemente simples del juego amoroso son distintas. En primer lugar, tanto si se trata de bacterias unicelulares como si son multicelulares, todas ellas se reproducen en cualquier caso sin ninguna intervención del sexo. Es más, la mayoría de bacterias existentes en la Naturaleza —por no decir todas ellas— son organismos multicelulares. En segundo lugar, cuando las bacterias se implican en el sexo se comportan de forma promiscua. Todas ellas pueden aparearse en cierto modo (dar y recibir genes) con bacterias de linajes completamente diferentes. Y toda esta cópula y todo este intercambio de genes involucran recombinación genética, pero no reproducción.
Y ésa es la cuestión. Para comprender el origen de las especies debemos antes ponernos de acuerdo sobre qué son, y en eso no somos nada coherentes. Los zoólogos y los botánicos se muestran normalmente satisfechos con la norma general de que los miembros de una misma especie producen descendencia fértil, mientras que los de especies distintas no. Pero con demasiada frecuencia, botánicos, zoólogos y otros biólogos asignan alegremente organismos a tal o cual especie basándose en la morfología, en la apariencia y el comportamiento de los cuerpos. Los bacteriólogos pueden utilizar la morfología únicamente en el caso de las bacterias más grandes, más visibles o más distintivas, como las mixobacterias, las espiroquetas y muchas cianobacterias. En los demás casos emplean una regla general distinta: cuando dos bacterias comparten el 85 por ciento de sus rasgos mensurables, se considera que pertenecen a la misma especie. Los bacteriólogos en activo son conscientes de la arbitrariedad de esta definición, habida cuenta de la rapidez con la que las bacterias cambian sus rasgos. Según esta norma general, las bacterias cambian todo el tiempo. Si comparten únicamente el 84 por ciento de sus rasgos mensurables, se considera que pertenecen a especies distintas. Colocadas en una nevera o en una incubadora, las bacterias pueden cambiar de «especie» en pocas semanas. Ningún zoólogo, botánico, micólogo o protistólogo estaña de acuerdo en que un mes en una nevera bastara para cambiar de una especie a otra.
Nuestra nueva definición de especie, en la que postulamos que los organismos con un mismo número y una misma clase de genomas integrados en común pertenecen a la misma especie, depende del reconocimiento de que todos los organismos nucleados son organismos compuestos. Todos ellos son el producto de simbiontes integrados. La idea consiste en que, para que dos organismos (individuos) sean miembros de la misma especie, tienen forzosamente que estar compuestos exactamente por el mismo conjunto de simbiontes. Puesto que las bacterias no están formadas por medio de la integración de simbiontes, en su universo no puede haber especies. La ventaja de nuestro análisis estriba en que todos los organismos nucleados son asignables a alguna especie, tanto si se implican en relaciones sexuales como si no. Si dos protoctistas comparten el mismo conjunto de genomas, bacterianos u otros —es decir, si comparten los mismos antepasados, si están construidos por conjuntos únicos de genes—, pertenecen por definición a la misma especie. Lo mismo puede aplicarse a los hongos, los animales o las plantas.
La «eucariosis» se refiere a la propia obra, al origen evolutivo de las primeras células nucleadas. Las primeras eucariotas fueron protoctistas. Los protistas —los miembros más diminutos del reino protoctista— evolucionaron por medio de la adquisición e integración de antepasados bacterianos específicos. Los primeros protoctistas deben haber sido protistas anaeróbicos en entornos libres de oxígeno. Antes de las eucariotas, es decir, antes de que evolucionara cualquier protista, el mundo era anóxico y enteramente bacteriano. No existía especie alguna en el eón Arqueozoico. Las primeras eucariotas, los primeros protoctistas, fueron las primeras especies sobre el planeta, aunque no —ojo al matiz— los primeros organismos. La adquisición e integración de genomas ajenos por primera vez no tuvo como resultado únicamente las primeras eucariotas, sino también la aparición de las primeras especies en la historia de la vida.
En la medida en que las relaciones entre bacterias con capacidades distintas se volvían irreversiblemente íntimas, estos complejos bacterianos acabaron generando la célula protoctista. Las eucariotas más pequeñas, los primeros protoctistas, condujeron a los antepasados microbianos de todas las formas de vida mayores.
Muchos de los protoctistas más pequeños, protistas por definición, viven tan felices sin nada de oxígeno. De hecho, este gas es para ellos un veneno letal instantáneo. Entre estos seres que evitan el oxígeno están los arqueoprotistas, los «protistas amitocondriados». Este filum de centenares de especies vive en hábitats tales como los intestinos de mamíferos e insectos, o en lodos ricos en sulfuro. Opinamos que estos protistas amitocondriados son representantes vivos de los antepasados de todos los organismos nucleados (ver figura 9.1). No pueden metabolizar el oxígeno ni reproducirse sexualmente, pero constituyen claramente especies. Por ejemplo, las dos especies nadadoras halladas en las termitas comedoras de madera seca —Metacoronympha kirbyii y Trichonimpha ampia— pueden distinguirse fácil y coherentemente.
Si aceptamos la idea de que las especies se originaron en las eucariotas, antes de la aparición de la respiración del oxígeno o de la meiosis (la clase de división celular implicada en la fertilización sexual), podremos resolver el dilema de Darwin sobre el origen de las especies desde el principio del fenómeno mismo de especiación. Una vez que nos ponemos de acuerdo en que tanto las especies como el proceso de especiación no existen en el mundo de las bacterias, las especies estables conocidas pueden ser nombradas, identificadas y clasificadas.

Figura 9.1 Metacoronympha y Trichonimpha, protistas digestores de madera en las termitas.
Existen bacterias con millones de formas distintas. Algunas fotosintetizan, muchas convierten pequeños compuestos orgánicos en miríadas de otros más complejos. Algunas acumulan hierro para producir imanes en sus propios cuerpos. Otras nadan hacia la luz o lejos de los ácidos. El mundo de las bacterias posee prácticamente todos los talentos que asociamos con la vida animal y vegetal. Las nuevas especies no aparecen cuando los organismos heredan «características adquiridas», sino cuando adquieren otros genomas, organismos completos con sus propias series enteras de genes, que determinan las características en cuestión. Jean-Baptiste Lamarck estaba en lo cierto: la herencia de características adquiridas ocurre, pero tan sólo cuando dichas características están determinadas por los genomas de organismos adquirióles. Charles Darwin también tenía razón: las nuevas especies aparecen cuando los organismos nuevamente adquiridos son integrados, dejan descendencia, y son perpetuados por la selección natural. El origen simbiogenético de las especies se basa firmemente en el trabajo de ambos, Darwin y Lamarck.
§. El Thiodendron de Dubinina y la sangre de Dennis
A pesar de que muchos estén en desacuerdo con nuestras conclusiones, los capítulos precedentes se basan en ciencia bien establecida. Relataremos ahora una historia peculiar en la que la ciencia está en progreso y en la que todos los detalles no están aún verificados.
¿Cómo evolucionó el núcleo, esta característica definitoria de las células de todos los organismos eucariotas? Probablemente no por mutación. El origen de la célula que constituye el antecesor común de toda la vida especiada tuvo, con toda seguridad, que involucrar «consorcios de bacterias», bacterias viviendo con y dentro de otras bacterias.
Érase una vez, hace unos 2.000 millones de años, en el eón Proterozoico, un organismo denominado Thiodendron latens medraba en zonas costeras de todo el mundo. Todavía se le puede encontrar hoy en día aquí y allá. (Ver en la tabla 9.1) un resumen de las divisiones en «tiempo geológico», la escala temporal completa.) Este Thiodendron, que tiene la apariencia de masas blancas de filamentos y huele a azufre, fue estudiado por el ecólogo microbiano ruso S. A. Perfiliev a mediados del XX. De hecho, fue este científico quien le puso por nombre Thio (azufre) dendron (árbol) latens (escondido), a la vista de sus células filamentosas, repletas de bolas amarillas y repartidas a intervalos regulares a lo largo de su cuerpo.

Según registró Perfiliev, de las células bacterianas filamentosas brotaban frecuentemente pequeñas células mótiles, nadadores flagelados que participaban en la producción del sulfuro de hidrógeno. Resumiendo, Perfiliev insistió en que el Thiodendron, como muchas otras bacterias tales como la Caulobacter y la Chondrococcus, acumulaban una historia vital dilatada y compleja, en la que los filamentos se alternaban con los organismos unicelulares flagelados. Sin embargo, y a diferencia de los otros dos géneros, el Thiodendron transformaba el azufre del ion SO=4 en la forma elemental (S), en forma de tiosulfato (HSO−4), o en su estado reducido (H2S). Pero ante la presencia de oxígeno, este organismo transforma el azufre disponible en el estado oxidado SO=4. La forma de azufre que el organismo produce depende, pues, de su disponibilidad de oxígeno.
Perfiliev murió antes de que se generalizaran las potentes técnicas del microscopio electrónico de corte fino. Le sucedió su discípula G. A. Dubinina quien, en el año 2002 y hacia el final de su carrera como microbióloga, seguía aún trabajando en el Instituto de Ecología Microbiana de la Universidad de Moscú.
Dubinina tenía interés por desvelar, mediante la alta ampliación, las etapas de la historia vital del desarrollo del Thiodendron latens de Perfiliev. Cosechó sus especímenes en los mismos lugares en que lo había hecho su maestro, en el mar Blanco por encima de San Petersburgo y también en los baños de barro del balneario de Staraya, en el lago Nijni, cerca de Moscú. Durante varios años, Dubinina y sus compañeros buscaron una descripción moderna del Thiodendron, basada en el análisis por medio del microscopio electrónico y en ensayos fisiológicos de producción de sulfuro, así como en otros trabajos de laboratorio. En líneas generales, tanto la propia Dubinina como varios estudiantes y diversas investigaciones postdoctorales confirmaron la descripción original de Perfiliev. Pero imaginémonos su sorpresa cuando, al tratar de cultivar Thiodendron en el laboratorio, colocaron las filamentosas masas sulfurosas bajo condiciones de anoxia completa y los filamentos, aunque no sus bastoncillos nadadores, se transformaron en hebras largas y enjutas. Los bastoncitos nadadores, identificados como Desulfobacter, la conocida bacteria reductora del azufre, continuaron nadando, manteniendo su aspecto y produciendo H2S como de costumbre. Los largos filamentos, en cambio, se fraccionaron en hebras helicoidales más cortas, indistinguibles de las espiroquetas típicas. Para que este cambio se produjera, era necesario retirar del medio absolutamente todo el oxígeno. Expuestos a una minúscula parte de oxígeno, los filamentos seguían oxidando el azufre elemental, que depositaban en el interior de sus células. Si la cantidad de oxígeno presente aumentaba, las espiroquetas se enderezaban. Sus células seguían siendo muy finas, pero se hacían cada vez más largas. Incapaces de dividirse, simplemente se alargaban. Tan sólo tras sobrevivir largo tiempo en su estado alargado acababan muriendo, presumiblemente debido a la exposición al oxígeno.
El grupo de Dubinia siguió trabajando, descubriendo al menos otros cuatro grandes grupos de Thiodendron marino, compuestos todos ellos por bacterias nadadoras reductoras de azufre muy parecidas y espiroquetas ligeramente distintas. Por ejemplo, en el océano Pacífico, en la isla Yankich de la bahía de Kraternaya, o en Papúa y Nueva Guinea, se encontró al Thiodendron en matas blanco azuladas de veinte milímetros de grosor. Al ser colocadas en el medio adecuado, sin oxígeno, todas las muestras produjeron terceras poblaciones de espiroquetas independientes. Los autores invalidaron el nombre de Thiodendron al comprobar que no se trataba de una única bacteria con un ciclo vital complejo, sino que habían conseguido aislar dos tipos de bacterias distintas: un bastoncito corto nadador que producía sulfuro de hidrógeno y una espiroqueta formadora de filamentos que medraba en presencia del sulfuro de hidrógeno que ella misma no podía producir. Las asociaciones estables entre bacterias completamente distintas se conocen bajo el nombre de «consorcios bacterianos». Como señalaron Dubinina y sus colegas, se había dado la denominación única de Thiodendron a lo que en realidad era un consorcio entre dos bacterias. El paso siguiente consistió en describir a cada uno de los miembros de ese consorcio. La primera de ellas, corta y con flagelos, es la Desulfobacter, la productora de sulfuro de hidrógeno que degrada el azúcar a medida que respira azufre y lo convierte en sulfuro. La otra, la Spirochaeta oxidante del sulfuro, lo convierte en glóbulos de azufre que almacena en su cuerpo. La espiroqueta vive de la energía y el alimento que extrae de las largas moléculas de celulosa procedentes de las paredes celulares de las algas en proceso de degradación, que son fragmentadas por las enzimas de celulasa en un azúcar denominado celobiosa. Muchos caracoles, hongos y bacterias costeras producen enzimas de celulasa. Las espiroquetas formadoras de filamentos viven, como casi todas, de la celobiosa, proporcionando al mismo tiempo subproductos de la fragmentación del azúcar a su socia reductora de azufre.
El trabajo de Dubinina fue publicado por primera vez en inglés en el año 1993, en la publicación rusa científica Mikrobiologiya, en donde el equipo de Dubinina señalaba que las espiroquetas pueden claramente ser geoquímicamente importantes para la transformación del azufre en áreas del litoral marítimo. Tan sólo después de recibir de la profesora Dubinina un paquete de fotografías nos dimos cuenta al momento de que habíamos visto a la Thiodendron numerosas veces, sólo que siempre la habíamos tomado por una bacteria costera ordinaria oxidante del oxígeno, como la Thiothrix, la Vitrioscilla o la Beggiatoa, todas las cuales tienden a crecer en forma de masas blancuzcas (figura 9.2).

Figura 9.2 Thiodendron latens en cultivo pobre en oxígeno.
Durante muchos años, nuestro compañero en la Universidad de Massachusetts Dennis Searcy ha venido defendiendo que el «citoplasma básico» de todas las células nucleadas habría evolucionado a partir de un microbio libre, sin paredes, comparable a la Thermoplasma acidophilum que cultiva en su laboratorio. Se trata de un organismo limitadamente mótil. A diferencia de la Desulfobacter, no tiene flagelos, ni siquiera está rodeado por una membrana celular. Puesto que el citoplasma no está limitado por ninguna pared rígida, la Thermoplasma cambia de forma con facilidad. Dennis y su equipo han recopilado pruebas de que este microbio producen proteínas de «motilidad». Estas proteínas son la base del fenómeno del movimiento. Permiten que las células se extiendan lentamente sobre los glóbulos de sulfuro que Dennis les ha proporcionado como medio en el laboratorio. La principal diferencia entre la Thermoplasma y la Desulfobacter, miembro del consorcio de Dubinina, estriba en que el organismo de Dennis vive en agua dulce caliente. Se hiela a temperatura ambiente, no soporta el agua salada y crece en acideces de un pH 2,5 más o menos, muy inferior al pH 8,0 de las aguas en las que fuera descubierto el Thiodendron. No obstante, la Thermoplasma acidophilum produce grandes cantidades de sulfato de hidrógeno. De hecho, le encanta vivir colgada de glóbulos de azufre elemental, alejada del oxígeno. Se pega a la superficie de esos glóbulos, e incluso los envuelve con su cuerpo. A diferencia de la Desulfobacter, clasificada como eubacteria, la Thermoplasma acidophilum pertenece al subreino arqueobacteriano de los procariotas. Contrariamente a casi todas las demás bacterias, produce proteínas similares a la histona con las que envuelve su ADN. Dennis sospecha que estas proteínas protegen su ADN del fuerte ácido y del elevado calor que, de otro modo, degradarían rápidamente su genoma por «hidrólisis ácida». El Thiodendron, por su parte, no necesita tales medidas extremas, habida cuenta de que este consorcio amante del azufre vive bañado por las aguas frías y neutras de los océanos.
Pero, ¿de qué modo se relacionan estas bacterias sulfurosas con la historia de las primeras eucariotas?
Dennis Searcy, quienes aquí escribimos y tal vez algún que otro converso más, sugerimos que el primer consorcio que se convertiría en la quimera que, a su vez, se convertiría en la primera eucariota, fue una asociación parecida al Thiodendron, Postulamos que una arqueobacteria del tipo de la Thiodendron productora de sulfuro de hidrógeno (más que una Desulfobacter) y una eubacteria gramnegativa, un organismo muy parecido a la Spirochaeta de nuestros días, ávida del sulfuro de hidrógeno para protegerse del oxígeno, formaron una alianza. Sin duda tuvo que haber mucho forcejeo y mucha destrucción antes de que la asociación se hiciera permanente. La selección natural por la natación rápida y la obtención de alimento forzó a ambos organismos a una estrecha asociación. Juntos degradaron azúcares y otros carbohidratos bajo condiciones anóxicas, ricas en azufre. Probablemente, la asociación prosperó durante milenios en aguas de dulces a saladas y de tibias a calientes, manteniendo la Thermoplasma la misma producción de sulfuro de hidrógeno que su socia Spirochaeta oxidaba de nuevo en azufre elemental.
Dean Soulia, un alumno de Searcy, se propuso producir un consorcio del tipo Thiodendron en el laboratorio, utilizando para ello el Thermoplasma acidophilum de Searcy y espiroquetas procedentes de nuestras colecciones de campo. En la primavera del 2002 el trabajo seguía en marcha. No tenemos noticias sobre si ha tenido éxito. La idea consiste en colaborar con Antonio Lazcano y Arturo Becerra de la Universidad Autónoma de México (UNAM), junto con el laboratorio del profesor Searcy. Hasta ahora, Soulia ha empleado los métodos que utilizara Louis Pasteur para cultivar organismos anaeróbicos en el laboratorio. Imita las reglas de selección natural descritas por Charles Darwin, colocando a sus organismos en agua caliente y ácida, desprovista de oxígeno, de modo que tan sólo los más «aptos» prosperen y tengan descendencia. El objetivo consiste en obtener un consorcio estable de Spirochaeta y Thermoplasma en un medio creado en el laboratorio.
Tan convencido está Dennis Searcy de que todas las eucariotas llevan consigo el legado de sus antepasados, que decidió someter su idea a la comprobación en el laboratorio. Buscó para ello células en las que pudiera estudiar la capacidad residual para producir sulfuro de hidrógeno al suministrarles azufre elemental. Finalmente optó por las células de su propia sangre, porque las células sanguíneas rojas de los mamíferos carecen tanto de núcleo como de mitocondrias. También se parecen a los termoplastos en otro aspecto: como todas las células animales, nunca tienen paredes celulares. De este modo —razonó Searcy— no podría confundirse con otros orgánulos celulares y podría centrar su atención en verificar la capacidad del propio citoplasma eucariótico para metabolizar azufre, capacidad que nadie tenía el menor motivo de esperar que ese citoplasma tuviera. No tan sólo descubrió Searcy que su propio citoplasma genera copiosas cantidades de sulfuro de hidrógeno cuando se le suministra azufre elemental, sino que extendió su observación a los cuatro reinos eucarióticos. Suministró azufre a levaduras y mohos y demostró que también los hongos producen sulfuro de hidrógeno (H2S). El sulfuro tenía que ser retirado en cuanto era producido, puesto que tendía a acumularse en forma de residuo tóxico, pero una vez tomada esta precaución, Searcy pudo demostrar fácilmente que todas las células nucleadas, sean éstas vegetales, animales, fúngicas o protoctistas, producen sulfuro de hidrógeno. Naturalmente, no podía retirar de estas células el núcleo y las mitocondrias para comprobar directamente la capacidad del citoplasma, como había hecho con las células rojas de su propia sangre, pero aún así, el resultado de su trabajo resulta convincente. Searcy mantiene (y nosotros le apoyamos) que nuestras células remotamente antepasadas comenzaron como anaerobios, en medios ricos en azufre. Aún recuerdan sus orígenes porque su medio original forma parte del metabolismo. El consorcio original podría perfectamente haber sido el de una Thermoplasma con una Spirochaeta, unidas por la oxidación y la reducción de azufre, que acabaron permanentemente pegadas y cada vez más integradas. La célula eucariótica se formó por alianza simbiótica, por medio de la adquisición de genomas bacterianos muy distintos entre sí.
El siguiente eslabón de la cadena —creemos— es la quimera de Radhey Gupta.
§. La quimera de Gupta
El profesor Radhey Gupta ya no conduce. Sus antebrazos han quedado tan desgastados con su trabajo en el ordenador, que ya no se fía de tomar el volante. Formado en el prestigioso Tata Institute of Science, en las hermosas colinas de las afueras de Bombay, Gupta llegó a Canadá en los 80 como profesor auxiliar de bioquímica de la Escuela de Medicina de la Universidad McMaster de Hamilton, Ontario. Su interés se centra en descifrar las relaciones entre formas de vida microbianas, en cuyo esfuerzo su principal herramienta consiste en la comparación entre las largas cadenas de moléculas que toda célula precisa para vivir: las proteínas y los ácidos nucleicos. Gupta centra principalmente su atención sobre las largas series de residuos de aminoácidos en las proteínas, básicamente de las bacterias. Tras una detallada comparación entre las secuencias proteicas de unos 300 tipos de bacterias distintas con proteínas parecidas, procedentes de protistas anóxicos, Gupta comenzó a indagar en la historia de la propia célula protista. Al igual que a nosotros, le interesan particularmente las células nucleadas que carecen de mitocondrias. Opinamos que algunas de estas protistas descienden de antepasados que nunca las tuvieron.
Escritor prolífico, Gupta es autor de numerosos trabajos, en los que se incluyen resultados y opiniones que contradicen la corriente principal en microbiología. Esta corriente científica general insiste, por ejemplo, en que todos los eucariotas evolucionaron directamente a partir de arqueobacterias. No habría estado involucrada ninguna clase de simbiosis o de simbiogénesis. Cal Woese, profesor de microbiología en la Universidad de Illinois, Urbana, y su equipo de colaboradores, han llegado incluso a rebautizar como «arqueas» a las células arqueo bacterianas, negando así su naturaleza bacteriana y elevando el grupo de las «arqueas» a un estatus paralelo al de otras bacterias y de todas las eucariotas, e insistiendo en la existencia de tres grandes grupos de formas de vida. Gupta no está de acuerdo con eso. Todas las eucariotas —argumenta—, incluso las que carecen de mitocondrias en alguna etapa de su vida, tienen un pedigrí dual, dos clases distintas de antepasados. Todas las células nucleadas contienen algunas secuencias proteicas que se parecen a las de las arqueobacterias (como la Thermoplasma) y otras secuencias proteicas que recuerdan a las de las eubacterias (como las espiroquetas). Gupta insiste en que no hay razón alguna para suponer que el antepasado eubacteriano de la célula nucleada fuera más parecido a una espiroqueta que a otra eubacteria cualquiera (aunque no está de acuerdo con nosotros acerca de qué grupo eubacteriano fue el antepasado de todas las eucariotas no dispone, hoy por hoy, de alternativa válida a la Spirochaeta). Con todo, le corresponde a Gupta en exclusiva el mérito de la utilización del término «quimera», así como de la afirmación de que todas las eucariotas son quimeras. Las quimeras eran bestias míticas de la antigua Grecia, con cabeza de león, cuerpo de camero y partes de cuerpos de otros animales. Eran compuestos de partes reconocibles de orígenes diversos. Gupta opina que esta descripción encaja con las células cuyas secuencias proteicas estudia. Todas las eucariotas —afirma—, incluyendo a las que carecen tanto de mitocondrias como de cloroplastos, y viven sin oxígeno o sin luz, son quiméricas. Las células que hoy consideramos como parientes cercanos a las primeras eucariotas son compuestas, tienen antecedentes tanto eubacterianos como arqueo bacterianos.
Aunque, como bioquímico que es, Gupta no utilice los términos «simbiosis» y «socio simbiótico», eso es precisamente lo que está describiendo a nivel molecular. Su quimera se formó cuando dos simbiontes —uno eubacteriano y el otro arqueobacteriano— se fundieron en una unión permanente. Todos estamos de acuerdo en que uno de los simbiontes fue una eubacteria (fuera ésta una espiroqueta, como la socia del Thiodendron, u otra), así como en que el otro simbionte fue una arqueobacteria (fuera ésta una productora de sulfuro, como la socia del Thermoplasma, u otra). Creemos —aunque no necesariamente lo crea Gupta— que la quimera tuvo que ser una buena nadadora. Por ello opinamos que la espiroqueta es un antepasado probable, habida cuenta de que está provista de propulsión propia. Las espiroquetas que aportaron motilidad al interior de las células nucleadas, propulsaban a su termoplasma asociado hacia aguas dulces, ácidas y ricas en azufre. El termoplasma, a su vez, con su producción de sulfuro de hidrógeno (H 2S) altamente reducido, que «barre» el oxígeno, protegía a la espiroqueta de este gas que, en aquel momento de la historia del planeta (hace unos 2.500 millones de años), se estaba convirtiendo en una toxina medioambiental cada vez más importante.
A medida que la ágil espiroqueta nadadora se iba vinculando cada vez más a su Thermoplasma amante del azufre, evolucionó la célula compuesta, la quimera. En su interior evolucionó un nuevo sistema de orgánulos, denominado cariomastigonte, en respuesta al imperativo de la integración simbiótica. En la historia de la vida sobre el planeta, las asociaciones que mantienen unidas a células distintas en alianzas permanentes, han evolucionado en numerosas ocasiones. Pero aun aceptando la idea del consorcio bacteriano (Thermoplasma y Spirochaeta), ¿cómo imaginar la transición de aquella quimera bacteriana hasta convertirse en un protista genuino, con su núcleo bien desarrollado? La clave está en la estructura misma de esta célula, bien conocida desde principios del siglo XX. El cariomastigonte, el núcleo, su conector proteináceo con el cilio o «látigo» (undulipodio), y el propio undulipodio, comprenden un sistema de orgánulos presente en muchas células y habitualmente ignorado. Su importancia evolutiva no ha sido reconocida hasta hace poco, y gracias a nuestro colega Michael Dolan, anteriormente estudiante de doctorado en la Universidad de Massachusetts. Dolan explica claramente por qué y cómo las primeras células eucarióticas contenían el cariomastigonte con su «conector nuclear». Esa parte del cariomastigonte es una cadena proteica que liga al orgánulo de motilidad —la anterior espiroqueta eubacteriana— al resto de la célula. En opinión de Radhey Gupta y Dennis Searcy, el resto de la célula era anteriormente una arqueobacteria independiente. Searcy, que señala el modo como el ADN está ligado a las proteínas, así como el modo en que sus células se mueven y se adhieren al azufre, declara, basándose en estos y otros argumentos, que el antepasado arqueobacteriano se parecía a la actual Thermoplasma. Estamos de acuerdo.
Tal como lo vemos, el propio núcleo envuelto en membrana evolucionó a partir de una fusión del ADN de la quimera. La unión entre los ADNs de la Thermoplasma y la Spirochaeta, otrora independientes, ocurrió como cualquier otro apareamiento entre bacterias: con la bien conocida transferencia de ADN de una procariota a la otra. La arqueobacteria Thermoplasma y la eubacteria Spirochaeta, al estilo típico de recombinación bacteriana, unieron sus ADNs creando un único genoma. Las células dentro de células proliferaron membranas. El nuevo núcleo, que definía a la nueva quimera, estaba rodeado por membranas y unido, desde el principio, al orgánulo de motilidad, la anterior espiroqueta. Ésta evolucionó hasta convertirse en el undulipodio, una estructura con muchos alias: cilios, flagelos eucarióticos, colas de esperma. Esta estructura celular móvil presenta un patrón característico y bien conocido de túbulos minúsculos [9(2)+2] sobre cuyo pasado común todos parecen estar de acuerdo. El sistema de orgánulos (núcleo, conector nuclear, undulipodio compuesto por cinetosomas y sus tallos axonémicos) fue nombrado «cario mastigonte» (de karyo = semilla y mastigont = látigo) por quienes lo vieron por primera vez. En 1933, el microscopista alemán Robert Janicki lo incluyó en la descripción de sus diminutos protistas mótiles, pero no sería hasta que Harold Kirby (1900-1952), zoólogo de la Universidad de California y presidente del Departamento de Zoología de Berkeley hasta su muerte, trabajara sobre el cariomastigonte, cuando llegaríamos a comprender lo que hace realmente.
Creemos ahora que comprendemos cómo evolucionó el núcleo, principalmente gracias al trabajo pionero de Kirby. El núcleo se originó como parte del cariomastigonte, pero luego quedó en libertad. La razón por la que creemos comprender los orígenes del núcleo consiste en que algunos de los organismos hoy en día vivos se comportan del mismo modo en que creemos que lo hicieron sus antepasados. Debemos reconstruir la historia evolutiva a partir de indicios vivos que consideremos representativos. La novedad evolutiva consistente en la célula nucleada se comprende mejor cuando se la considera como el producto histórico de asociaciones y simbiosis, de fusiones bacterianas de ADN cuyos productos (proteínas, ARN, lípidos) interactuaron, generando estructuras emergentes. Las mutaciones aleatorias tan sólo refinan y alteran el cambio a nivel de especie, pero no lo provocan. Las simbiosis dilatadas condujeron a la simbiogénesis, al origen de nuevos orgánulos, nuevos sistemas organulares, nuevos tejidos, nuevos órganos, nuevos organismos y nuevas especies. La simbiogénesis —la herencia de genomas adquiridos—, principalmente entre bacterias u otros microbios, constituye la fuente mayoritaria de innovación evolutiva. La selección natural dirige la evolución a través de la propagación y eliminación de lo que ya tiene. Sin embargo, la simbiogénesis —como en el caso de la fusión de una arqueobacteria y una eubacteria en la quimera de Gupta— constituye el principal proveedor de materia prima, entre la que la selección natural se encarga de elegir.
§. La liberación del núcleo
El cariomastigonte, ese extraño sistema organular común a los protistas y al esperma, se compone de un undulipodio (al menos un cinetosoma-centriolo [9(3)+0] y su correspondiente tallo [9(2)+2], el axonema) conectado al núcleo por el rizoplasto o «conector nuclear». Ver una versión simplificada en la figura 9.3.
Figura 9.3 Un cariomastigonte (izquierda) comparado con un acariomastigonte (derecha). (Protistas seleccionados útiles para la reconstrucción de la evolución del cariomastigonte, aquí descrita).
Seguimos necesitando de esos términos para comprender, en primer lugar, cómo surgió esta clase de células. Janicki, Harold Kirby, D. H. Wenrich y sus coetáneos crearon estas etiquetas para describir las estructuras móviles que distinguían en las células vivas y en sus preparaciones tintadas. Aquellos investigadores las consideraban como requisitos previos a la comprensión de la individualidad, la reproducción, la sexualidad, el género y otros procesos fundamentales de la vida de nuestros antepasados unicelulares. Tan sólo ahora, casi un siglo después, podemos encontrar el sentido para la evolución del cariomastigonte, del acariomastigonte y de la paradesmosa, de esa plétora de túbulos, fibras, membranas, orgánulos y sistemas organulares completos de humildes protistas.
La cuestión fundamental es nuestra idea de que el cariomastigonte (cinetosoma/centríolo-conector nuclear-núcleo) constituye una estructura «emergente» que se formó por primera vez en la primera de las eucariotas. ¿Por qué? El sistema organular fue seleccionado con el objetivo de prevenir la pérdida del látigo celular, del orgánulo nadador que la quimera necesita, tanto para evitar con rapidez el oxígeno como para obtener alimento. Tuvo que existir una fenomenal presión evolutiva para que los genes de la arqueobacteria original (como la Thermoplasma) se adhirieran a los de su eubacteria asociada (como la Spirochaeta) tan fuertemente, que ni siquiera la muerte pudiera separarlos. En resumen, el cariomastigonte constituye el legado del imperativo original, para los miembros de toda asociación simbiótica, de permanecer unidos y de ser heredados juntos. El cariomastigonte comenzó con su bolsa membranosa en la que sus dos socios bacterianos depositaron sus genes: el núcleo. En algunos linajes, el cariomastigonte se atrofió y dejó tras de sí a cinetosomas proliferando en el borde de la célula. En otros el cariomastigonte entero se reprodujo mucho más deprisa que su citoplasma, generando nuevas especies de células con ocho, treinta o incluso mil cariomastigontes.
Figura 9.4. Snyderella tabogae (muchos núcleos libres y aún más acariomastigontes).
En otros linajes celulares protistas, como el que desembocaría en la Snyderella (figura 9.4), el núcleo se desvinculó de su cariomastigonte original, permitiendo la proliferación de acariomastigontes. En este linaje, los acariomastigontes —cada uno con su correspondiente undulipodio— proporcionaron presumiblemente mayor poder natatorio a sus células portadoras. El resultado fue la evolución de una célula gigante cargada de núcleos libres y de aún más acariomastigontes. Nuestro relato evolutivo, en el que el cariomastigonte (el núcleo amarrado) precede al núcleo liberado, queda reforzado con la observación de laGyronympha. Este género de multinucleados, pariente de la Snyderella, contiene numerosos acariomastigontes y núcleos libres (sin atar, excepto en la división celular). Casi como si se hubiera olvidado de desprenderse de ellos, la Gyronympha retiene también unos pocos cariomastigontes (núcleos amarrados). Una vez que se ha conseguido evolucionar buenos planos corporales, ¿para qué prescindir de ellos?
Investigadores pioneros, como Janicki, Kirby y Lemuel Roscoe Cleveland, estudiaron lo que ellos denominaban animales primitivos o protozoos, para tratar de averiguar cómo había evolucionado la vida animal. Describieron al cariomastigonte con su núcleo vinculado, así como los cambios que se producían a medida que aquellas «células animales simples» crecían y se dividían. Primero se reproducía el «centro de la célula», el cinetosoma/centríolos en la base del undulipodio. Dos se convertían en cuatro y cuatro en ocho. La mayoría de células de algún interés estaban relacionadas con la Trichomona vaginalis, causa de un picor vaginal muy molesto. Éstas tendían a presentar cuatro cinetosomas/centríolos en la base de cuatro tallos. En la división celular, estos cuatro se convertían a menudo en ocho, con una fina fibra que separaba los viejos de los nuevos. Esta fibra se prolongaba en una línea que los primeros biólogos denominaron «paradesmosa» (de para = junto a, a lo largo de, y desmosa = conector). El modo en que el cariomastigonte se convierte en la paradesmosa cuando la célula se divide fue bellamente demostrado por Kirby. Los núcleos, a veces centenares en una sola célula, adoptaban forma de pesa de gimnasia. Se estrechaban para duplicarse. Luego, el núcleo viajaba hasta la paradesmosa y se montaba en ella a medida que ésta se iba estirando hasta que, finalmente, ésta transportaba a cada uno de los dos núcleos descendientes a ambos lados opuestos de la célula, que iban a convertirse en dos. En ausencia de la paradesmosa alongada, los núcleos se dividían pero no se separaban. Eso hizo que Kirby considerara a la paradesmosa como una especie de huso mitótico. Al igual que sucede con el huso mitótico de los animales, la paradesmosa duplica sus cinetosomas/centríolos, crece, se alarga y se disuelve. Como el huso, también la paradesmosa actúa separando sus núcleos descendientes durante la división celular.
El microscopio electrónico confirma la visión de Kirby. Demuestra que la paradesmosa está compuesta por microtúbulos de una media de 240 micrómetros de diámetro (24 nanómetros), exactamente igual que el huso. La estructura consiste en una delgada y primera versión del huso mitótico. Consideramos que el cariomastigonte de la célula que no se divide y la paradesmosa que forma durante la división, son herencias de la fusión simbiótica entre arqueobacterias y eubacterias. La paradesmosa es la estructura proteica que garantiza que los genomas de las bacterias asociadas, antes independientes, sean heredados juntos. Fue precisamente esta distinción la que convirtió en permanente a la alianza entre eubacterias y arqueobacterias.
Parte IV
Consorcios
Ladrones de nematocistosDonald I. Williamson, de la Universidad de Liverpool, ha pasado toda su vida profesional en el Laboratorio Marino de Port Erin de la isla de Man. Allí, desde el aislamiento de su banco marino, ha formulado «un proceso a saltos en la evolución animal, que opera con independencia de la acumulación de mutaciones y de la selección».
Un calamar luminoso
Los olénidos de Fortey
Resumida a su esencia, lo que la declaración de Williamson nos dice es que los cambios evolutivos importantes en los invertebrados —y casi todos los animales lo son— emanan de la herencia de genomas adquiridos. Williamson argumenta que los genomas que determinan las formas animales larvales difieren de las que determinan su forma adulta y no proceden del mundo microbiano, sino que su origen debe buscarse en otros animales. A diferencia de los genomas ajenos llamativamente coloreados —habitualmente en verde—, que delatan la presencia de bacterias fotosintéticas en algas y plantas, la transferencia entre animales de genomas acuosos e incoloros que defiende Williamson tiende a ser pasada por alto.
Williamson explica que el origen de las larvas —formas inmaduras de insectos, estrellas de mar y de tantos otros moradores de los acantilados y los lodos marinos— habla de extrañas y arcanas simbiosis tan integradas, que de ella quedan tan sólo oscuros indicios. «Discuto la presuposición, ampliamente aceptada, de que las larvas y sus correspondientes adultos han evolucionado siempre dentro de un mismo linaje, y presento mi hipótesis alternativa de transferencia larval —escribía en el 2001 en el Zoological Journal ofthe Linnean Society—. [Mi] hipótesis nació de la convicción de que la evolución enteramente dentro de linajes separados resulta inadecuada para explicar la distribución de tipos larvales en el reino animal, así como de que los métodos de metamorfosis que vinculan las fases sucesivas del desarrollo no podrían haber evolucionado simplemente por selección natural y mutaciones aleatorias. Subrayo, sin embargo, que no propongo la transferencia larval como substituto de la selección natural. Adultos y larvas han evolucionado gradualmente por medio de “descendencia con modificación” pero, de forma sobrepuesta a este proceso, genomas enteros han sido transferidos por hibridación».
Williamson postula que los encuentros sexuales, ya sea por fertilización externa (huevos puestos y fertilizados en el agua) o interna (entrada de esperma en el cuerpo de la hembra con ayuda del pene del macho), ocurrieron en ocasiones entre individuos de clase y filum muy distintos con un éxito espectacular en algunas ocasiones. Estos apareamientos exitosos entre animales muy distantemente emparentados se dieron con muy poca frecuencia, unas treinta o cincuenta veces en 541 millones de años. Esto significa que un apareamiento exitoso y fértil ocurre aproximadamente tan sólo una vez cada 10 millones de años. Williamson toma la sucesión, dentro de un mismo proceso de desarrollo, de dos o más formas corporales distintas (larvas de una clase que se convierten en adultos de otra) como prueba directa de la presencia de genomas distintos pero integrados (heterogenomas).
Los datos que llevaron a Williamson a esta sugerencia radical ocupan mucho espacio en la bibliografía de la zoología marina. Adultos extremadamente diferentes (erizos, estrellas) comparten tipos de larva casi idénticos (denominados plúteos), mientras que adultos próximamente emparentados (una segunda especie de erizo de mar u otra estrella) se desarrollan a partir de larvas completamente distintas (bipinnarias). Williamson cita a la gran zoóloga Libby Hyman, de la Universidad de Chicago, y a su obra de cuarenta volúmenes sobre los cincuenta filia de animales, publicada en el año 1940. Al describir a los miembros del filum de los celentéreos, Hyman escribió: «Hidroides a menudo próximamente emparentados producen medusas muy diferentes y viceversa, de modo que las medusas pueden ser clasificadas en familias distintas a partir del hidroide del que proceden».
Los encuentros sexuales fértiles entre animales adultos de linajes muy distintos han sido presenciados al menos en una ocasión: machos de erizo marino (Echinus esculentus, animal del filum de los equinodermos) fueron vistos por Williamson fertilizando huevos de cordado. (El de los cordados es nuestro supetfilum. En la actualidad, los animales conocidos con cráneo están ubicados en el filum de los craneados, dentro del supetfilumde los cordados. Este incluye toda la «megafauna carismática»; tigres, caimanes, tortugas de las Galápagos, elefantes, tiburones, cóndores gigantes, etc.). El cordado madre cuyos huevos fueron fertilizados por erizos de mar era el Ascidia mentula. No sólo los huevos fertilizados sobrevivieron a aquella extraña unión, sino que se convirtieron en larvas completamente fértiles, en formas inmaduras denominadas plúteos. Sin embargo, y a diferencia de los erizos de mar normales, la mayoría de estas larvas nacidas de los huevos anómalos de cordado no consiguieron desarrollar órganos adultos normales. En lugar de brazos larvales produjeron esferoides: se convirtieron en animales redondeados, cada uno de ellos con un disco adhesivo. Este comportamiento es desconocido en el desarrollo normal de los huevos de Echinus fertilizados. El Ascidia, en cambio, produce siempre discos adhesivos, que le permiten al animal adulto adherirse a la roca u otro sustrato sólido, tras superar la fase larval en forma de renacuajo. Algunos de estos híbridos entre filia sobrevivieron hasta noventa días después de salir del huevo. Unas pocas larvas desarrollaron órganos rudimentarios típicos de su padre erizo y, transcurrido un plazo de entre treinta y siete a cincuenta días, se convirtieron en erizos de aspecto enteramente normal, como si su herencia cordada hubiera desaparecido definitivamente. Cuatro años más tarde, los tres erizos supervivientes produjeron huevos. Williamson fertilizó estos huevos de híbrido con esperma de erizo marino en libertad. De esa fecundación resultaron larvas plúteas normales. Incluso su ADN ribosómico y mitocóndrico era típico del Echinus. Aparentemente, en los animales que habían desarrollado la forma de erizo adulto únicamente había sobrevivido el genoma equinodermo paterno. Sin embargo, en la mayoría de descendientes esferoides, había prevalecido el genoma de la Ascidia méntula. Este extraño cruce de filia es análogo a los experimentos ordinarios de cruce de Mendel dentro de una misma especie, donde el rasgo que aparece en el híbrido no refleja el gen recesivo, sino el dominante. Sólo que, en el caso del cruce de filia entre el Echinus y el Ascidia, un genoma haploide completo, cruzado con otro genoma haploide completamente distinto, revela cuál de los genomas completos domina en primer lugar: el del erizo de mar. El desarrollo posterior se produce de tal modo que la «dominancia genómica» se relaciona con el estado en la historia de la vida.
En el caso de este cruce entre erizo marino macho y hembra de Ascidia, el genoma haploide paterno predomina y el materno desaparece. Aunque se intentó el cruce genómico inverso, es decir, de un padre Ascidia con una madre Echinus, nunca prosperó. Una gran oportunidad para estudiar el apareamiento ilegítimo, es decir, la hibridación entre distintos yifia, espera al genetista zoólogo (¿o tal vez deberíamos decir genomicista?) animoso que se decida a emprender esta línea de investigación.
La realidad del cruce genómico está clara para Don Williamson. Ha sido testigo de la hibridación entre animales que no solamente no pertenecen a la misma especie, sino tampoco al mismo género, a la misma familia, al mismo orden, a la misma clase, ni siquiera al mismo filum. La gran inferencia consiste en que, hace unos 225 millones de años, ocurrieron fusiones de este tipo entre equinodermos tales como ciertas estrellas de mar. Algunas de ellas carecen, aún hoy, de estado larval, desarrollándose como adultas directamente desde el huevo. Otras en cambio, como atestigua Williamson, adquirieron genomas completamente nuevos por medio de la fusión. Fueron adquiridas algunas clases de nadadores de plancton adultos, y éstos dieron pie a formas larvales en estrellas de mar que antes carecían de ellas. De hecho y según Williamson, todos los equinodermos carecían de larvas hasta inmediatamente después de la mayor extinción de especies de la historia de la vida, la permotriásica que ocurrió hace 225 millones de años. Al comenzar el Mesozoico, los equinodermos supervivientes fértiles de diversas clases adquirieron y retuvieron de forma independiente a adultos con los que se habían apareado ilegítimamente. Como Williamson ha demostrado, esta tendencia hacia la promiscuidad y los gustos exóticos no ha desaparecido del todo. Los equinodermos hoy existentes podrían muy bien seguir siendo capaces de adquirir larvas de urocordados como la Ascidia, con su simpática forma de renacuajo del que brotan discos adhesivos cuando llega el momento de pasar a la vida adulta.
Williamson ha ampliado su teoría. En su libro de 1992 sobre larvas incongruentes, teorizaba que tan sólo determinadas especies, distribuidas en ocho filia, habrían adquirido sus fases larvales por transferencia. Ahora, en cambio, asegura que todas las especies productoras de larvas, incluso las orugas y otros animales terrestres, adquirieron genomas ajenos en algún momento de su historia. Williamson cita itinerarios de desarrollo altamente complejos desde larvas múltiples hasta la fase adulta, por ejemplo en la gamba decápoda de la familia de los sergéstidos. Este crustáceo metamorfosea cuatro veces durante su desarrollo desde el huevo hasta un animal adulto con diez patas. Las cuatro larvas son planctónicas, lo que dificulta la comprensión sobre cómo se acumularon las mutaciones que las generaron en un medio uniforme (los protoctistas y los animales bentónicos, es decir, que viven en el suelo del océano, tienden a ser mucho más variados que sus parientes planctónicos, tanto en el presente como en el registro fósil). Numerosas gambas decápodas mueren durante la metamorfosis, lo que denota una intensa selección en los híbridos, pero las que sobreviven se transforman de huevo a naupliomorfo, a plenocárido, a misidáceo, a mastigopoido y a adulto sergéstido. Esta plétora de transformaciones larvales en una misma gamba decápoda le sugiere a Williamson la persistente presencia del pasado. La gamba sergéstida —sugiere— adquirió, integró y puso a trabajar al menos cuatro genomas intactos. Para Williamson, la herencia de estos genomas adquiridos —que no mutaciones aleatorias— determina el éxito evolutivo de esta gamba hasta nuestros días.
En sus seminarios, Williamson gusta de mostrar una transparencia del conocido árbol de las familias animales de Ernst Haeckel, del siglo XIX. Este árbol expresa únicamente diversificación, ramas que se subdividen en otras. No se describe en él ninguna fusión y mucho menos ningún cruce entre animales asignados a distintos filia. Por supuesto —admite Williamson—, Haeckel demostró ser un científico esforzado y muy creativo, « ¿Pero por qué —se pregunta Williamson—, habida cuenta de que Haeckel diseñó su filogénesis basándose en datos recopilados antes de 1880, debería seguir estando hoy en lo cierto?». La pregunta planea en la sala junto con la transparencia de la zoología haeckeliana. «Échenle un buen vistazo a esta filogenia —le dice Williamson a su reducida y atenta audiencia— porque está a punto de extinguirse».
Tras estudiar a las larvas en el mar, en el laboratorio y en la biblioteca, Williamson ha acumulado numerosas razones para dudar de que animales marinos de tipos ampliamente distintos, hayan evolucionado por mutación aleatoria a partir de larvas extraordinariamente parecidas, cuando no idénticas. Williamson rechaza el árbol de Haeckel y el estilo de pensamiento neodarwinista. En su lugar postula la adquisición, integración y herencia de genomas entre supervivientes, como la norma general en la evolución de animales con larvas anómalas. Las fuentes de las diversas larvas —concluye Williamson— son varias. Incluyen rotíferos, que dieron lugar a las larvas trocóforas de anélidos y moluscos. Los onicóforos adultos, conocidos como «gusanos de terciopelo», se parecen sospechosamente a las orugas. Tal vez —sugiere Williamson— proporcionaron sus genomas larvales a lepidópteros (polillas y mariposas), a algunos neurópteros (como las moscas-escorpión) y a la mayoría de himenópteros (abejas, avispas) asignados al suborden de sínfitos (ver figura 10.1).Presumiblemente los demás subórdenes de himenópteros, con larvas completamente distintas, no eligieron las mismas parejas. Miembros de distintos órdenes de insectos adquirieron genomas onicóforos por hibridación.
Los renacuajos expanden el catálogo de Williamson. Los larváceos (apendicularios) de nuestro superfilum de los cordados, clasificados con los ascidios como urocordados (cordados con cola), no metamorfosean en absoluto, sino que permanecen como renacuajos toda su vida, incluyendo la fase reproductora de la misma. Las larvas renacuajo —argumenta Williamson— fueron adquiridas cuando larváceos adultos consiguieron donar con éxito sus genomas, probablemente por hibridación, a los ascidios, que presentan larvas peculiares en forma de pez. En general y según Williamson, los metazoos adultos que se desarrollaron directamente a partir del huevo precedieron en la evolución a cualquier forma larval. Esta opinión es verificable: de ser correcta, el registro fósil debería revelar, de forma rutinaria, que los adultos precedieron a las larvas. Es más, puesto que las mismas clases de larvas se han interpuesto entre diferentes linajes en distintos momentos del eón Fanerozoico, algunas larvas deberían haber sido adquiridas por linajes ajenos incluso muy recientemente.
Figura 10.1 Las «larvas transferidas» de Williamson.
§. Ladrones de nematocistos
Examinaremos tan sólo dos ejemplos más de herencia de genomas adquiridos en animales marinos. En ambos casos, los detalles de la fusión nos ayudarán a comprender el potencial de la evolución a saltos, de la especiación rampante y de la apariencia de taxones genuinamente novedosos. Observaremos los hidramedusoides del filum cnidarios y, dentro de los moluscos, a los cefalópodos que se desarrollan directamente a partir del huevo y brillan en la oscuridad: Euprymna scolopes.
Al filum de los cnidarios pertenecen las hidras de agua dulce, la fisalia o carabela portuguesa, los sifonóforos de las chimeneas de las profundidades marinas y las anémonas en forma de flor de los charcos de marea. Todos los miembros de esta antigua división animal tienen células urticantes activadas por disparadores, que dan muy mala fama a todo el filum. Estas células urticantes, denominadas nematocistos, son complicados sistemas de orgánulos empaquetados dentro de células especializadas denominadas cnidocistos. Durante muchos años, los ctenóforos estuvieron agrupados junto con los hidromedúseos en el filum único de los cnidarios, fundamentalmente por la semejanza de sus respectivos nematocistos. Puesto que los ctenóforos, con sus largas extensiones repletas de cilios densamente apretujados, tienen muy poco más en común con los hidromedúseos de generación alterna, aparte de sus células urticantes, los dos filia han sido separados de nuevo en celentéreos (hidromedusoides) y ctenóforos.
Elie Metchinikoff, descubridor ruso de las células macrófagas comedoras de bacterias en la sangre de los cangrejos, solía pasar sus veranos en el sur de Italia, en el estrecho de Messina, que separa a Reggio di Calabria de Sicilia. En la última parte del siglo XIX había allí una excelente base naval construida por la armada del zar. Como muchos otros biólogos prosoviéticos, Metchinikoff podía llevar a cabo sus investigaciones en un enclave vacacional europeo. Antes de la Gran Guerra, muchos científicos rusos o de otros países del Norte contribuyeron enormemente a las investigaciones biológicas marinas desde esta clase de emplazamientos veraniegos privilegiados.
La brillante contribución a la ciencia de Metchinikoff, derivada principalmente de su estudio de las células fagocíticas del cangrejo, incluye no tan sólo la teoría celular de inmunidad, sino también su insistencia sobre la sorprendente falta de correlación en la clasificación de los hidrozoos. Estos celentéreos se parecen a la Hydra aunque, sin embargo, su historial incluye también una fase de «cuenco invertido», al estilo de las medusas. La fase hidroide de algunos de ellos se correlaciona perfectamente con la fase medusa del mismo animal, mientras que en otros las dos etapas no tienen la menor relación entre sí: la medusa que se les parece pertenece a una especie completamente distinta. Williamson sugiere que esta incongruencia entre clasificación adulta y larval no es más que otro caso de adquisición de un conjunto de genomas (los hidroides) por otro conjunto (los medusoides). La alternancia de generaciones hidroides con medusoides es enteramente diploidea: en todos los historiales, tanto del hidroide con tentáculos, como de su alternativa medusoide de cuenco invertido, son diploides. En otras palabras, todas las células de su cuerpo tienen dos conjuntos de cromosomas. Williamson afirma que su idea de que estas historias personales no siempre estuvieron unidas ayuda a explicar por qué algunos hidrozoos carecen totalmente de estado medusoide, mientras que otras «hidromedusas» no tienen ninguno. Explicaría también por qué, en un mismo individuo celentéreo (con ambas etapas en su desarrollo) se puede comprobar que los medusoides se mueven por separado de los hidroides. Este comportamiento independiente implica separación de conexiones nerviosas, como si estuvieran presentes simultáneamente, en el mismo individuo, dos sistemas nerviosos completamente distintos. «Las dos formas corporales se originaron como los genomas de dos animales originariamente independientes», escribió Williamson. Mientras que la construcción de una forma corporal única de hidroide o medusoide implica el habitual genoma único diploide, la ocurrencia de dos formas corporales distintas durante el desarrollo, particularmente con movimientos musculares no relacionados, es considerada por Williamson como un argumento en defensa de la fusión de dos genomas anteriormente separados.
Mientras que esta tesis acerca del doble origen de los celentéreos es aún debatible, la probabilidad de que las células urticantes —los nematocistos— de los celentéreos fueran otrora organismos vivos autónomos, cuyos genomas fueron adquiridos independientemente por ctenóforos e hidromedusoides, nos parece muy plausible. El candidato a urticante independiente adquirido es un microsporidio, bien descrito por Stan Shostak y Victor Kolluri en 1995.
En estos «animales gelatinosos» la especiación, e incluso el origen de taxones más inclusivos, parece entenderse mejor como un efecto más de la herencia de genomas adquiridos.
Las superficies de los hidroides que carecen de etapas medusoides, como los tubulados, están típicamente recubiertas de urticantes. El término nematocisto se refiere a la célula urticante allí donde resida. Los nematocistos ovales tienen púas aguzadas con un tubo envenenado enrollado en su interior. Cada célula está armada, lista para disparar. Cuando el disparador es estimulado mecánicamente, los dardos con su veneno son proyectados para desgracia, y a menudo muerte, de la presa o del predador potencial. La mayoría de criaturas marinas se cuidan mucho de molestar a los bien protegidos hidroides.
Sin embargo, las babosas desnudas, moluscos sin cáscara denominados nudibranquios eólidos, se alimentan sin peligro de hidroides armados de células urticantes. En realidad casi no comen otra cosa. A menudo su color se asemeja al de su alimento. Los miembros eólidos de esta familia de babosas hacen ostentación de los bienes robados o, mejor dicho, de los genes robados. Estos caracoles sin cáscara ingieren partes de los hidroides y digieren lo que no necesitan guardar. Devoran todo el cuerpo de su presa menos las células urticantes. Luego desactivan estas células de modo que nunca lleguen a dispararse, algo así como tragarse una pistola cargada. Los nematocistos completos viajan por el tracto digestivo como cualquier otro alimento, sólo que de algún modo resisten a la digestión.

Figura 10.2 La nudibranquia Hermissenda, una ladrona de nematocistos.
La subclase de moluscos opistobranquios, dentro de los que se ubica el suborden de los eólidos, parece particularmente adaptada para sustraer genomas ajenos. Los ascoglossa, otro orden de moluscos gasterópodos, tienden a ser de color verde. Roban sus genomas a fotosintetizadores en lugar de nematocistos. La observación de ochenta y seis especies probó que un 82% eran ladrones de genomas teñidos de verde. Natural de las marismas saladas desde el sur de Florida hasta Nueva Escocia, la Elysia chlorotica retiene plástidos fotosintéticos de la Vaucheria litorea. Si le falta el alimento externo, esta babosa puede vivir foto autotróficamente durante meses. La Elysia tímida se alimenta exclusivamente de jóvenes Caulerpa, un alga grande, verde y monocelular, reteniendo sus cloroplastos. Según el investigador que la estudió, la supervivencia de esta Elysia «depende por completo de su adaptación al ciclo vital del alga clorofícea Acetabularia acetabulum». La Elysia tuca toleras las algas calcificadas y recolecta sus cloroplastos de partes no calcificadas de la Halimeda incrassata. Nos encontramos aquí de nuevo con la correlación entre especies reconocidas y la clase de genomas que retienen.
§. Un calamar luminoso
La adquisición de un genoma ajeno —el de la gammaproteobacteria Vibrio fischeri— convierte a un pequeño calamar en una estrella reluciente del espectáculo nocturno (figura 10.3) en este caso, la relación genética es poderosa: ningún calamar de esa especie carece de su complemento completo de bacterias bioluminiscentes (luz fría).

Figura 10.3 El pequeño calamar luminoso Euphryma scoleps
Los calamares nacen de huevos sin las bacterias. Excretan cada día el 90% de sus habitantes bacterianos. Lo que realmente heredan de sus progenitores son unos genes animales que controlan el desarrollo de un curioso tejido: un enorme proyector de dos lóbulos, alojado en el vientre del calamar, una corteza epitelial especial, un músculo translúcido modificado y un reflector oscuro que dirige la luz fría. Con el paso del tiempo evolutivo, el cuerpo de este calamar ha sufrido una transformación masiva para convertirse en receptáculo de sus moradores bacterianos. Sin embargo, si no está presente en el medio la cepa adecuada de bacterias vivas, potencialmente huéspedes, todo el órgano luminoso del animal queda sin formarse.
Margarett McFall-Ngai y Ned Ruby mantienen una colonia de entre diez y doce parejas de calamares en su laboratorio de la Estación Marina de Kewalo de la Universidad de Hawaii. Estas parejas cautivas ponen unos 30.000 huevos al año. En caso de que ocurra algún percance en los tanques, McFall-Ngai y Ruby no tienen más que salir a la playa por la puerta del Pacific Biomedical Research Center y coger ejemplares directamente del océano.
A diferencia de muchos de los moluscos de Williamson, el Euprymna scoleps carece de etapas larvales. Los embriones se desarrollan durante unos veinte días en las aguas del océano y, sin cuidados parentales, los calamares salen del huevo al anochecer. Todos los recién nacidos exhiben órganos luminosos tan sólo inmaduros.
El órgano luminiscente de la cría de calamar está recubierto de cilios. Un anillo de cilios batientes empuja el agua hacia una serie de poros, tres a cada lado del órgano. Cada poro lleva a un conducto y de allí a una cripta donde se alojan las bacterias. El órgano luminoso del calamar está así dispuesto, con su anillo de cilios, para sus residentes bacterianos, desde el momento en que el animal sale del huevo. A las doce horas, la respiración del calamar y la actividad de los cilios han capturado suficientes bacterias, que se alinean en el espacio intercelular. La población de bacterias comienza entonces a crecer por división hasta llenar el órgano. Tan sólo la adecuada cepa de Vibrio fischeri puebla el órgano luminoso del Euprymna scoleps. De algún modo, el calamar parece ser más apto que el mejor de los microbiólogos para identificar y cultivar las bacterias adecuadas a partir de la mezcla confusa de las aguas oceánicas. Unas doce horas después de haberse mudado a su nuevo hogar, las bacterias envían un mensaje de muerte masivo e irreversible al tejido del calamar que las alberga. Las células epiteliales de la superficie ciliada mueren bajo esta orden. Unos cuatro días después de salir del huevo, no queda sobre el calamar huella alguna de su anterior dispositivo de «bienvenida». La señal es sutil e indirecta. Las bacterias tienen que penetrar en la cripta nadando. Deben inducir un incremento cuádruple en el volumen de las células epiteliales de la superficie del calamar. La cantidad de microvello o pequeñas proyecciones de las superficies superiores de la cripta tiene que incrementarse espectacularmente. Si los simbiontes bacterianos son eliminados mediante calor o antibióticos, el juego termina de inmediato. El órgano luminoso vuelve a su estado anterior, parecido al que tenía el calamar al salir del huevo. Los genes bacterianos que codifican la proteína «adhesina» de la membrana celular de la bacteria parecen ser los promotores del mantenimiento del órgano luminiscente.
¿Por qué están los sistemas del calamar y de su bacteria residente tan bien sintonizados para el desarrollo de ese enorme órgano luminoso? Aparentemente, la razón es la «contrailuminación», una especie de camuflaje. Desde abajo, tanto los predadores como las presas potenciales del calamar luminoso ven lo que parece ser el cielo de la noche, en lugar del apetitoso vientre del calamar. La luz ventral generada por las bacterias es de la misma intensidad, color y distribución angular que la de la Luna y las estrellas. Mientras que durante el día el calamar relleno de bacterias duerme en la arena del fondo, en sus correrías nocturnas en busca de alimento revolotea iluminado a contraluz por sus simbiontes bacterianos. La experimentación ha demostrado que los calamares desprovistos de su apéndice luminoso son presa fácil de sus predadores naturales. Aunque estos animales desprotegidos puedan sobrevivir sin problemas en el laboratorio, en su medio natural no puede existir ningún Euprymna scoleps sin la compañía de sus simbiontes.
§. Los olénidos de Fortey
Aún disponiendo de organismos vivos para su estudio, la inferencia de la especiación y la evolución de taxones superiores por simbiogénesis está cargada de dificultades. Esta dificultad es aún mayor cuando se trata de detectar la simbiogénesis en el registro fósil cámbrico. Sin embargo, eso es precisamente lo que ha hecho el paleontólogo británico Richard Fortey. Los trilobites de la familia olénidos dejaron abundantes caparazones (recubrimientos corporales) en los esquistos negros de finales del Cámbrico y del Ordoviciense (figura 10.4). Los trilobites, una clase dentro del filum de los artrópodos, tenían patas con articulaciones pero, a diferencia de sus parientes, los insectos y los crustáceos actuales, no estaban recubiertos por quitina. Sus caparazones estaban formados principalmente por carbonato cálcico. Estos animales llenaron los mares del Paleozoico de una fascinante variedad. Algunos eran tan pequeños como moscas, mientras que otros tenían el tamaño de una langosta grande; algunos apenas se movían, otros eran asesinos rápidos y feroces. Hacia el período Pérmico, hace 260 millones de años, todos se habían extinguido, tras haber permanecido en escena durante unos gloriosos 300 millones de años, cien veces más que nuestra especie, al menos de momento.

Figura 10.4 de Richard Fortey: Olenus en sus yacimientos «No-hay-nadie-en-casa».
Prácticamente todos los trilobites exhiben una placa ventral denominada hipostoma, que actuaba durante la manipulación del alimento. El primer y más simple olénido, el mismísimo Olenus, tenía una hipostoma normal. En una de las familias, la de los pelturinos, esta placa bucal se degeneró completamente. Abundan las mudas articuladas intactas. Éste y otros indicadores sugieren que los olénidos prosperaron en un entorno de aguas tranquilas. Es más, sus mudas indican que se trataba de animales más bien aplanados, con músculos axiales débiles. Al parecer vivían en lugares donde las corrientes de fondo eran escasas. Los caparazones olénidos se encuentran piritizados, es decir que el carbonato cálcico que los formó fue posteriormente embebido en sulfuro de hierro —pirita—, también conocido como “el oro de los tontos”. La pirita se forma inmediatamente cuando el sulfuro (gas H2S) entra en contacto con agua rica en hierro. Fortey imagina que los trilobites olénidos debían nadar en aguas tranquilas, ricas en sulfuro y pobres en oxígeno.
A diferencia de la mayoría de los demás trilobites, los olénidos tienen cutículas extremadamente finas. Exhiben estructuras lamelares en forma de peine denominadas «salidas». Algunos paleontólogos interpretan estas estructuras como «ramas branquiales», como protecciones de las extensiones (branquias) que «respiran» o intercambian gas disuelto en los peces con agallas parecidas. A no dudar, estas extensiones incrementaron en gran medida la superficie de contacto con aquel agua rica en sulfuro. Fortey, que trabaja desde muy pronto por la mañana hasta bien entrada la noche con estos fósiles en el departamento de Historia Natural del Museo Británico en Kensington, Londres, sugiere que todos los trilobites olénidos tenían simbiontes bacterianos. Como sucede con las actuales lombrices tubiformes de las chimeneas submarinas (vestiminíferas), diversos géneros de anélidos (lombrices segmentadas) y moluscos bivalvos con asociaciones simbióticas extremadamente desarrolladas, en el caso de los trilobites podemos tomar los cambios morfológicos como indicadores de simbiotrofia. Los animales vivos que cultivan tales simbiontes bacterianos en sus tejidos muestran signos distintivos de su asociación. Como en el caso de los fósiles de Fortey, muchos han modificado su boca u otras partes de su aparato digestivo. Algunos portan trofosomas (órganos para almacenar bacterias) hipertrofiados o lámelas branquiales. Por otro lado, los animales asociados a bacterias tienden a reproducirse en grandes números.
Algunos olénidos tienen bolsas de incubación espectacularmente grandes. Fortey sugiere que las larvas (crías) de trilobite eran protegidas dentro de tales bolsas, hasta que hubieran cultivado suficientes simbiontes bacterianos fijadores del dióxido de carbono y oxidantes del sulfuro, como para poder vivir de forma independiente. En la Gran Cantera de Andrarum, Suecia —los «esquistos de alumbre» del Cámbrico superior, hace 505 millones de años—, la que se conoce como zona Olenus ha sido investigada con detalle por Euan N. K. Clarkson, de la Universidad de Edimburgo. A lo largo de un metro y medio de una sección de esquisto, puede verse una clara correlación entre la presencia de pirita —sulfuro de hierro— y dos especies de olénidos: O. truncatus y O. wahlenbergi. Cuando la pirita desaparece, los olénidos también, y les sustituye un fósil ostrácodo. Cabe deducir que el Cyclotron, una especie de crustáceo marino, evitaba las aguas ricas en sulfuro y vivía allí donde el oxígeno hacía la vida soportable. Al parecer, dominaba allí donde podía respirar oxígeno. Los olénidos, en cambio, huían de cualquier fuga de este gas. Fortey cree que estos antiguos trilobites construyeron sus caparazones gracias a los favores autotróficos de las bacterias fijadoras de dióxido de carbono que vivían en su cuerpo. Los olénidos cultivaban bacterias, del mismo modo que la Staurojenina de Andrew Wier (ver capítulo 7) y los calamares hawaianos luminosos de Margaren McFall-Ngai y Ned Ruby. Las bacterias oxidantes del sulfuro fijan el carbono como lo hacen las plantas y las algas, pero no crecen en presencia de oxígeno gaseoso. En los olénidos de Fortey, estas bacterias pudieron incluso existir en cultivo puro dentro del cuerpo de los trilobites. De este modo, 500 millones de años antes de Louis Pasteur y sin siquiera la ventaja del lenguaje, los artrópodos marinos olénidos parecen haber desarrollado la capacidad de cultivar bacterias. Aquel arreglo conveniente les ahorró a los trilobites el engorro de buscar comida. Las bacterias oxidantes de sulfuro parecen haber servido bien al imperativo reproductor de los olénidos, puesto que este gran suborden generó sesenta y cinco géneros y tal vez un centenar de especies. Bajo aquellas condiciones de abundancia de azufre y escasez de oxígeno, los otros trilobites, carentes de simbiontes bacterianos, se habrían asfixiado y muerto de hambre.
Capítulo 11
Proclividades vegetales
El paraguas del pobre§. El paraguas del pobre
La vida secreta del Geosiphon
Nichos fijadores de nitrógeno
Las relaciones parasexuales fértiles con apareamientos extraños y teóricamente estériles han sucedido a lo largo de la historia de la vida entre organismos terrestres de diferentes especies… y siguen sucediendo hoy. La Gunnera manicata, una herbácea de hojas enormes, crece en la selva bajo las nubes que cubren los Andes. En Ecuador, a unos seis mil metros sobre el nivel del mar, esta planta, a la que se conoce popularmente como «paraguas del pobre», mantiene una asociación con la cianobacteria Nostoc (figura 11.1).Esta bacteria contribuye a distribuir nitrógeno, siempre escaso, al resto de miembros de la comunidad de la planta, infectando las glándulas de su tallo e introduciendo sus filamentos en túneles especiales a lo largo de los pecíolos de sus hojas. Allí, en colaboración con la planta, esta cianobacteria realiza una fijación simbiótica del nitrógeno dentro de sus heterocistos. La Gunnera no es, ni mucho menos, el único morador sedentario del planeta fertilizado de este modo por cianobacterias; bien al contrario, esta clase de acoplamientos es abundante.
Existen en la actualidad unas 10.000 cianobacterias distintas, identificables tan sólo por unos pocos expertos. Es probable que, antes de que evolucionara cualquier alga o planta, existieran muchos más tipos de cianobacterias que, lamentablemente y al igual que tantos otros organismos del pasado, hoy están extintas.

Figura 11.1 La Gunnera manicata y su cianobacteria simbionte.
Los 30 millones de especies de hongos, animales, plantas y protoctistas se vieron forzados a adquirir el nitrógeno de sus proteínas por otros medios. Algunos superaron su permanente deficiencia de nitrógeno uniéndose literalmente a bacterias fijadoras de nitrógeno y adquiriendo sus genomas de forma temporal o permanente. La consecuencia de este comportamiento condujo a nuevos taxones, nuevas especies, nuevos géneros y, en algunas ocasiones, a familias y clases nuevas. Describiremos aquí tan sólo algunas de las consiguientes novedades evolutivas para apoyar nuestra propuesta de que la innovación evolutiva no depende nunca exclusivamente de la acumulación de mutaciones aleatorias, ni siquiera de la duplicación de genes bien desarrollados. En los casos que aquí mencionaremos, el taxon en cuestión (plantas y hongos nuevos) evolucionó cuando el hongo o la planta adquirieron e integraron de forma estable, a tiempo parcial o completo, el genoma de una bacteria fijadora de nitrógeno.
§. La vida secreta del Geosiphon
El Geosiphon pyriforme, de unos pocos milímetros de altura, fue descubierto a principios del siglo XIX en Neustadt, Alemania, por un profesor de ciencias de educación secundaria, que recogió la «planta» en la tierra empapada de la ribera del río y la registró en el herbario con el nombre de «Botrydium». Estas diminutas vejigas en forma de pera, que suelen estar reunidas en grupos de a tres a diez, son hoy en día extremadamente difíciles de encontrar. Carecen de registro fósil y, de hecho, sólo han sido estudiadas en Alemania y en la Europa Central. Nadie las ha visto nunca ni en América del Norte o del Sur, ni en el Pacífico. Fritz von Wettstein, un excelente botánico, volvió a etiquetar y rebautizó el organismo en el registro herbario cuando se percató de su verdadera naturaleza. Ahora, tras muchos años de estudio tanto en el laboratorio como en el campo, D. Mollenhauer ha conseguido por fin desvelar la vida secreta del Geosiphon. Se trata de un organismo doble, parecido a un liquen, con dos tipos distintos de antepasados. Uno de ellos es un hongo que se parece al Endogone, al Glomus y a otros miembros bien conocidos del filum fúngico zigomicetes. El otro es la cianobacteria Nostoc, fijadora del nitrógeno. El hongo se vuelve verde, aumenta hasta un tamaño «monstruoso» (para un hongo) de unos cuarenta y cuatro milímetros y produce unas peculiares vejigas, desconocidas en ningún otro caso descrito por la literatura botánica.
Tanto el hongo como la Nostoc cambian radicalmente en la asociación. Fijan los dos gases atmosféricos de importancia para su cuerpos: el dióxido de carbono (vía fotosíntesis) y el nitrógeno (vía nitrogenasa). Estas útiles fijaciones ocurren únicamente en el organismo integrado «parecido a una planta». Bajo la influencia del hongo, los resbaladizos propágulos de la cianobacteria, denominados «hormogonias», quedan atrapados. Se hinchan y se detienen. Una vez que la inmovilización de la cianobacteria está asegurada, tiene lugar una transformación mágica. La pared celular del hongo se rompe y sus membranas se abren al exterior. En la brecha se forman luego una nueva pared y una nueva membrana. El hongo se traga las bacterias a través de la abertura y luego la cierra por completo. En una notable metamorfosis, la nueva asociación madura hasta convertirse en algo parecido a una planta. Las vesículas fijan activamente nitrógeno y carbono durante unas semanas hasta que, finalmente, los organismos se disocian y comienza de nuevo la extraña fertilización. Como género y especie, el Geosiphon se originó —como tantos otros— por simbiogénesis. Tanto desde el punto de vista evolutivo como del desarrollo, el Geosiphon es enteramente un producto de interacciones simbióticas.
§. Nichos fijadores de nitrógeno
Otros géneros y especies de plantas se originaron también simbiogenéticamente por la razón de costumbre: el hambre de nitrógeno. Los campos de arroz tienden a ser de un color verde brillante. Los manchones verdes que aparecen bajo los tallos del arroz parecen ser algas. Los tallos no parecen tener tronco, hojas ni raíces. Un examen cuidadoso demuestra que se trata de plantas; lo que parecen ser los talos aplanados de un alga son en realidad hojas. Estos moradores de los arrozales son diminutos helechos acuáticos. En bolsas de la parte posterior de sus hojas vive la Anabaena, otra especie de bacteria filamentosa adepta a la fijación de nitrógeno. Esta bacteria se desliza en la cavidad abierta de la hoja inmadura, que luego crece cerrando la apertura. Se han nombrado y estudiado seis especies distintas de Atolla. Unas células especiales de este helecho sirven de «estanterías» para mantener a las bacterias en su lugar.
Las cicadáceas son bien conocidas para cualquiera que visite Miami Beach u otros lugares de Florida en invierno. Parecen ananás gigantes caídas en el suelo. Muchas decoran las entradas de los hoteles en enormes macizos circulares. Son nativas de los trópicos de todo el globo, aunque abundan particularmente en Cuba y Sudáfrica. Las cicadáceas forman un taxón superior, un filum o división de plantas que incluye docenas de especies. Todas ellas son simbióticas con cianobacterias fijadoras de nitrógeno. No existe en la Naturaleza ninguna que carezca de sus genomas bacterianos cautivos. Sospechamos que la asociación cianobacteriana inició la proliferación de especies de las cicadáceas. Las cianobacterias no pueden encontrarse en las semillas de la planta, sino que penetran en el plantón en las primeras veinticuatro horas después de que germine. Las cicadáceas del tamaño de un árbol forman pequeños órganos verticales especializados en el suelo colindante, denominados «raíces coraloides». Un corte de estas raíces presenta un anillo verdeazulado, perfectamente integrado, que a la vista del inexperto podría resultar indistinguible del resto de las células de la planta. Ahí es donde residen las células procarióticas, adquiridas e integradas en las células de las raíces de la cicadácea, de las cianobacterias fijadoras de nitrógeno, que también en este caso son las Nostoc.
La plétora de plantas, árboles, hierbas, viñas, y demás leguminosas del Cretáceo, miembros todos ellos del orden Leguminosae, espectacularmente exitoso, evolucionó por simbiogénesis. La historia es demasiado larga, enrevesada y técnica para ser contada aquí. Basta con que, la próxima vez que te comas un guisante o una alubia, recuerdes que una de las razones de que este alimento rico en nitrógeno sea tan bueno para ti, estriba en que la planta de la que procede alberga en sus raíces un genoma bacteriano. La intimidad de la relación entre la leguminosa (incapaz por sí misma de utilizar el nitrógeno del aire) y la bacteria (que por sí misma tampoco fija el nitrógeno) es notable. Estos socios están integrados simultáneamente en los niveles genético, de productos genéticos, de genes, metabólico y conductual. La asociación entre las bacterias rizobiáceas del suelo y las pequeñas raíces filamentosas de la planta de la habichuela, que se prestan a su invasión natatoria en fila india, data al menos de 100 millones de años. Es probable que la simbiosis productiva se encuentre detrás de la proliferación y la especiación de esta maravillosa familia de plantas florecientes. Los fijadores de nitrógeno bacterianos no son fotosintetizadores. De hecho, en el caso de las legumbres dependen de los azúcares y otros productos de fotosíntesis de las plantas.
En el caso de las legumbres, los detalles de la adquisición de bacterias y su integración cíclica son bien conocidos, habida cuenta de que la industria trata de imitar a la Naturaleza. Las empresas agrícolas tratan de enseñar al maíz y al trigo (miembros de la familia de las herbáceas) a capturar los genomas de las bacterias rizobiáceas, con el objetivo de producir grano más rico en proteínas y más barato que otras fuentes de nitrógeno. Hasta la fecha no lo han conseguido. Las sutilezas de la historia de la integración entre leguminosas y rizobiáceas son abundantes. En caso de llegar a tener éxito estos esfuerzos industriales, las especies Zea (maíz), Hordeum (cebada) o Triticum (trigo), merecerían nombres de especie nuevos: Zeaazotogenica, Hordeum nitrogenicum o tal vez Triticum nutricium. De nuevo estaríamos asistiendo al origen de las especies no por mutación aleatoria, sino por adquisición genómica.
Capítulo 12
El baile de los cromosomas: la teoría de la fisión
La reproducción cinetócora como legado bacterianoNadie es más consciente de la importancia de los cromosomas que unos futuros padres, a la espera de los resultados de una amniocentesis. Esperan que se les confirme que su feto no tiene ni más ni menos que cuarenta y seis cromosomas, pulcramente alineados en veintitrés pares emparejados. Si desean un varón, sólo variará el último par (XY). La gran X no se corresponderá con la pequeña Y. Es ese cromosoma Y, que incluye muy pocos genes (principalmente para la formación de espermatozoides y de su capacidad para nadar), que determina el sexo masculino del futuro bebé. Todas las células de las niñas sanas tienen invariablemente cromosomas emparejados, incluyendo los cromosomas X, de los que niñas y mujeres tienen dos. La amniocentesis es esencialmente un análisis de cariotipo. Un axioma de la biología de los mamíferos reza que los miembros de la misma especie tienden a tener cariotipos idénticos, o al menos muy parecidos.
El cariotipo se define como el número total y la morfología del cromosoma completo de un animal o una planta. Las preparaciones carioptípicas proceden de células del cuerpo en proceso de división mitótica, a partir de las que pueden prepararse y guardarse preparaciones permanentes. Las células son fijadas (tratadas con compuestos químicos que mantienen su estructura discernible) y teñidas con tinturas cromosómicas, que permiten que los detalles de los cromosomas puedan ser tabulados y fotografiados. Los pares de cromosomas son entonces numerados y alineados según su tamaño, de mayor a menor. Las imágenes digitales o las fotografías reales de los cromosomas pueden ser interpretadas. Para ello, su calidad debe garantizar que pueda leerse el cariotipo. A menudo está en juego una nueva vida humana.
Los detalles del cariotipo han venido siendo recogidos en más de un millar de especies de mamíferos y centenares de especies de plantas desde principios del siglo XX. Puesto que los cariotipos son generalmente específicos para cada especie, el análisis cariológico es una herramienta de ayuda útil en la identificación de especies.
Si el gradualismo darwiniano es una explicación para los orígenes de las especies de animales y plantas, de ello se desprende que las especies próximamente emparentadas deberían tener cariotipos parecidos. No es así. Tomemos como ejemplo al ciervo asiático, especie del género Muntiacus. Estos ciervos se parecen mucho entre sí y, bajo comprobación, son aparentemente aptos para aparearse y producir descendencia. Las siete u ocho especies de Muntiacus van desde el este al oeste de Asia, abarcando un área de varios millones de kilómetros cuadrados. A pesar de que, indudablemente, no se dispone de todos los datos, el número de cromosomas de sus diploides (células corporales) varían desde un mínimo de tan sólo tres pares hasta un máximo de veintitrés.
Neil Todd, un recién licenciado en zoología de la Universidad de Harvard, desarrolló a principios de los años 70 su teoría sobre la fisión cariotípica, que relaciona la evolución de los cromosomas con la historia evolutiva de los mamíferos.
Como señalaron dos comentaristas, Todd le puso por nombre a su concepto Teoría de fisión cariotípica para «llamar la atención sobre su rechazo implícito al gradualismo darwiniano en la evolución cromosómica». Todd aplicó su análisis a tres grupos de animales: carnívoros cánidos, artiodáctilos (ovinos, vacunos, ciervos y demás ungulados de dos dedos) y (junto con el estudiante John Giusto) monos del viejo mundo y simios. Sus trabajos fueron ignorados o rechazados por los evolucionistas de la corriente mayoritaria. M. J. D. White, en su libro de 1973 titulado Citología animal y evolución, rechazó la tesis central de Todd con la siguiente observación: «suponer que todos los cromosomas de un cariotipo sigan ese mismo proceso [fisión en el centrómero-cinetócoro] de forma simultánea, equivale a creer en los milagros, lo cual no tiene cabida en la ciencia» (página 401). No resulta, pues, sorprendente que pocos zoólogos y evolucionistas se sintieran motivados para tomarse en serio el trabajo de Todd. A diferencia del profesor White, Todd carecía de laboratorio propio para el estudio de cromosomas. Tampoco era profesor titular ni autor de un libro ampliamente citado. De hecho, Neil Todd era —y sigue siendo— numismático y genealogista, toda una eminencia en escudos de tabernas irlandesas entre los siglos XVII y XIX, así como en genética gatuna.
Zoólogos y evolucionistas continuaron trabajando bajo las hipótesis, ampliamente respaldadas por White y otros neodarwinistas, de que los números de los cromosomas eran muy similares entre las especies emparentadas de mamíferos, y que las excepciones que Todd había señalado eran triviales. El mantra, ampliamente repetido, consistía en que el número de cromosomas en los mamíferos era elevado en las poblaciones antepasadas y fueron descendiendo gradualmente con el tiempo. La fusión de cromosomas, como ocurre en los humanos con síndrome de Down, sucedía de forma impredecible y daba cuenta de los cambios cromosómicos. El mensaje consistía en que Todd, creyente en los milagros, debía ser ignorado. A excepción de algunas publicaciones esporádicas, la historia entera de la fisión cariotípica permaneció en letargo hasta 1998.
Sin embargo, ha sido recobrada por razones que atañen la tesis de este libro. La fisión cariotípica en sí misma no es un proceso simbiogenético, pero sugerimos que está relacionada en última instancia con el origen simbiogenético de los cromosomas, como el propio Todd (2001) ha señalado.
Gracias al extraordinario desarrollo de los campos de la biología celular y del estudio de los cromosomas, al talento de una joven bióloga, Robin Kolnicki, que ha analizado los datos cromosómicos de los lémures, y al interés renovado del propio Todd por su gran idea, la situación ha cambiado radicalmente. La visión de Todd constituye la explicación mejor y más sencilla para los cariotipos mamíferos y el cambio cromosómico a lo largo del tiempo. La cosecha por parte de Kolnicki de abundantes pruebas procedentes de otro campo de la ciencia, demuestra por qué y cómo, sin necesidad de milagros, las ideas de Todd acerca de las radiaciones adaptativas resultan ser acertadas. «Radiación adaptativa» se refiere a la proliferación en la diversidad de especies en un momento dado del registro fósil, que aquí se correlaciona con cambios espectaculares en los números de los cromosomas de los mamíferos. Los investigadores noveles menos proclives a aceptar irreflexivamente la hipótesis de esos pequeños pasos graduales por los que se supone que procede la evolución, pueden aplicar cada vez más la visión Todd-Kolnicki a murciélagos, roedores, a otros mamíferos e incluso a pájaros y reptiles. Los orígenes de especies nuevas de mamíferos se correlacionan con el cambio cariotípico exactamente en el modo a saltos en que Todd anunció. Aquí tan sólo podemos revisar curiosamente las ideas básicas de esta visión. La literatura científica sobre ellas es aún limitada y manejable, habida cuenta de lo reciente de su nacimiento. Como sucede con otros aspectos del desprecio neodarwinista por la nueva evidencia, el debate ha comenzado vivamente.
Cuando, a principios de los años 30, comenzó a practicarse el análisis de cariotipo, espoleado por la teoría cromosómica de la herencia, numerosos científicos quedaron sorprendidos al encontrarse con que animales similares no tenían necesariamente cariotipos parecidos. La teoría de fisión de Todd postula un antepasado mamífero con un número diploide de catorce cromosomas «mediocéntricos» (el centrómero-cinetócoro se encuentra próximo al centro del cromosoma). La fisión completa de todos los cromosomas a la vez se toma como base teórica para explicar los brotes de especiación.
En poblaciones con todos los cromosomas grandes y mediocéntricos, la fisión generará un conjunto completo de cromosomas más pequeños y acrocéntricos (con el centrómero-cinetócoro hacia un extremo del cromosoma) y un número diploide doble del original. En el caso de los lémures, este número va desde veinte a setenta cromosomas. Los lémures, un suborden de primates nativos de Madagascar, se agrupan en cinco familias: lepilemúridos, daubentónidos, lemúridos (o eulémures), queirogálidos e índridos. Un mínimo de cuatro fases evolutivas, comenzando con el hipotético antepasado (probablemente parecido al actual Lepilemur ruficaudatus) con un número diploide de veinte, genera en principio cario-tipos en todas las especies vivas de lémures. Una primera fisión condujo a una gama de números diploides de entre veinte y treinta y ocho cromosomas. La siguiente fisión explica los lemúridos (de cuarenta y cuatro a sesenta y dos cromosomas) y los queirogálidos. En los antepasados de los índridos ocurrió una fisión secundaria distinta, así como otra fisión posterior independiente en los lepilemúridos. Quedan detalles por precisar, pero la teoría de la fisión reduce elegantemente la necesidad de centenares de explicaciones a medida para muy pocos acontecimientos de fisión seguidos por cambios más pequeños en los cromosomas de grupos reducidos de descendientes.
§. La reproducción cinetócora como legado bacteriano
Si los términos científicos te aburren, ahórrate esta parte del libro. Basta con que te digamos que la tendencia de los cromosomas a reproducirse, fraccionarse, reagruparse y cambiar según su propio plan refleja, probablemente, su vida pasada como entidades independientes. Los comportamientos de estas estructuras portadoras de genes influyen profundamente sobre nosotros, sobre los animales y sobre las plantas, cuyas células en crecimiento no pueden pasarse sin ellos. Esta sección analiza cómo la Teoría de la reproducción cinetócora explica, en términos biológicos novedosos y de un solo plumazo, la Teoría de fisión cariotípica de Todd, basándose en un único acontecimiento plausible y en sus consecuencias.
Los cinetócoro-centrómeros son los puntos de anclaje de cada cromosoma al huso mitótico, las pistas de microtúbulos sobre las que circulan los cromosomas para acceder a los extremos opuestos de la célula en fase de división. Ahora se sabe ya que se trata de estructuras dobles de proteínas (el cinetócoro) que se ligan a los microtúbulos del huso mitótico, manteniendo la continuidad del ADN (el centrómero) con el resto de los genes en un solo paquete. Cada vez que un cromosoma se reproduce, este paquete cinetócoro-centrómero/proteína-ADN se reproduce también. La reproducción cromosoma-cinetócoro-centrómero ocurre cada vez que la propia célula se reproduce.
El acontecimiento único plausible consiste en una reproducción extra del cinetócoro-centrómero seguida por la ruptura entre los pares de cinetócoros. Esta ronda extra y esta ruptura suceden simultáneamente en todos los cromosomas.
Por ejemplo, si sucediera en tus células productoras de esperma, ocurriría a la vez en todos tus veintitrés pares de cromosomas, que tienen sus centrómeros localizados medio céntricamente. Este sencillo acontecimiento plausible generaría una nueva célula con cuarenta y seis pares de cromosomas, ahora más cortos. En cada nuevo cromosoma, el cinetócoro-centrómero estaría ahora situado en un extremo. Dicho en la jerga del ramo, el suceso de fisión cariotípica convierte cuarenta y seis mediocéntricos en noventa y dos telocéntricos sin cambio significativo alguno en la secuencia de los genes (orden de los pares de bases del ADN), ningún cambio en la dosificación de genes (las ratios entre genes permanecen intactas) ningún cambio en la cantidad de ADN (la distribución de los genes en un número mayor de grupos de unión ocurre sin ninguna alteración en el ADN total) ni ningún otro cambio en el propio animal. La redistribución de cromosomas y la selección de sus agrupaciones después de la gran fisión son inevitables, habida cuenta de que, al aparearte, el esperma fisionado fertiliza un óvulo que, evidentemente, tendrá los normales veintitrés cromosomas mediocéntricos largos, en lugar de tus cuarenta y seis. ¿Por qué? Pues porque tanto la parte femenina de la pareja como sus células tendrán el número típico en nuestra especie.
La naturaleza ha ensayado este experimento muchas veces. Los animales fisionados y no fisionados son plenamente fértiles hasta que ocurre la redistribución de cromosomas (inversiones pericéntricas u otras al margen del ámbito de lo aquí expuesto). La reproducción cinetócora como explicación para la fisión significa una profundización crucial sobre los cambios (aceleración) en la secuencia temporal de producción de nuevos cinetócoro-centrómeros. Estos cambios en el ritmo, este retraso en la reproducción cromosómica en relación con la de cinetócoro-centrómeros, subyace en gran parte de la diversificación de especies de mamíferos. En determinados grupos de mamíferos estos cambios han sido demostrados con detalle para explicar el taxón: carnívoros, mamíferos artiodáctilos (ovino, vacuno, cérvidos), lémures, monos del viejo mundo y simios (chimpancés, orangutanes, gorilas, monos cercopi-técidos, babuinos y demás).
Los cinetócoro-centrómeros son puntos distinguibles en los cromosomas, regiones estrechadas donde se unen las fibras de los husos (microtúbulos) (figura 12.1). Los cinetócoro-centrómeros pertenecen a una clase de estructuras denominadas «centros organizadores de microtúbulos [18]», porque «capturan» microtúbulos y los atan al resto del huso mitótico. La producción de nuevos cinetócoro-centrómeros ocurre generalmente entre mitosas, cuando todo el ADN cromosómico restante también se está reproduciendo. Todo cuanto hace falta para que empiece a rodar la bola de la fisión cariotípica cuesta abajo hacia una nueva especie de mamíferos, es una ronda extra de síntesis de ADN cinetócoro-centrómero en relación con el resto de los cromosomas.
Figura 12.1 La Teoría de la Fisión Cariotípica (reproducción del centrómero-cinetócoro).
Escribe, «los simbiontes endocelulares tienden a reproducirse de forma desincronizada en relación con sus anfitriones, incluso en los casos de asociaciones simbióticas eubacterianas coevolucionadas como las de mitocondrias y plástidos. La teoría de la reproducción cinetócora (teoría de fisión cariotípica), en la que la “partición” del centrómero se entiende como una reproducción centrómera residual rápida de un genoma otrora ajeno (de eubacteria espiroqueta), se enteramente coherente con un concepto “simbiogenético”, y no de “filiación directa”, de la evolución de la célula eucariota».
Por supuesto, estamos de acuerdo. Y añadimos la visión de la especiación a saltos, discontinua, de Todd-Kolnicki a otras como la simbiogénesis y la transferencia larval de Williamson. Éstas son algunas de las formas como se crea la novedad evolutiva. Las especies se originan por herencia de genomas adquiridos, junto con sus rebeliones reproductivas dentro de los genomas anfitriones. Así es como proceden los taxones superiores tales como géneros, familias y demás. El proceso no es gradual, como tampoco pueden ser ajenos a él los animales, si tomamos en consideración estos «shocks postraumáticos» en la fisión de los cromosomas de los mamíferos.
Capítulo 13
Darwin de nuevo: las especies en el diálogo evolutivo
La idea fundamental consiste en que las características heredadas de importancia extraordinaria para la selección natural, tales como frutos, ojos, alas o habla, tienen siempre en común una dilatada historia precedente. El reconocimiento de los antecedentes eucarióticos, así como de la genética y el desarrollo de las eucariotas, convierte a lo que podría parecer intervención divina en evolución como de costumbre. La aparición evolutiva de ojos no es ninguna excepción. La capacidad para responder a la luz visible de forma específica, aparece en muchas moléculas comunes a todas las células. La capacidad foto sensitiva del tejido retinal humano (y de todos los demás vertebrados) es una propiedad de los bastoncillos y los conos de las células retinales. Químicamente, estas células contienen rodopsina, un complejo pigmento proteínico de color púrpura sensible a la luz. La componente opsina de la rodopsina es una proteína que cambia de un organismo a otro, pero sin dejar de compartir unas características comunes, y está presente en una gran variedad de animales, algunas bacterias y algunos protistas. Las verdaderas reacciones a la luz ocurren en la otra parte, más pequeña, de la molécula: la retinal. El nombre rodopsina procede del griego: rhodo, término griego para el color púrpura, se refiere a la parte retinal, mientras que opsina comparte la misma raíz optós que óptico, que se refiere al ojo. En una traducción poco rigurosa, el nombre de la molécula activa significaría pues proteína púrpura del ojo. La pequeña porción molecular que recibe directamente la luz data, en la historia de la vida, de mucho antes de que aparecieran los animales y las plantas. La rodopsina es visiblemente activa en todo el grupo de arqueobacterias denominado halófilas. Los procariotas, amantes de la sal, gustan de bañarse en la luz del Sol. Utilizan su rodopsina foto sensitiva para generar energía en forma de la omnipresente molécula ATP. Los fundamentos de la fotosensitividad están bien desarrollados en el mundo bacteriano.
Los bastoncillos (utilizados mayoritariamente en la visión nocturna) y los conos (para la visión diurna) son células alargadas, ricas en membranas, que se alinean en la retina de los animales. En nuestros ojos, éstas células presentan los habituales [9(3)+0] cinetosomas micro tubulares que sustentan el tallo microtubular, el axonema [9(2)+2] que, naturalmente, ya hemos estudiado al hablar de la motilidad intracelular. El bastoncillo no es más que un superdesarrollado pliegue interno paralelo de la membrana undulipodial (cilio). El cono es lo mismo, a diferencia únicamente de que las membranas plegadas están en capas concéntricas en lugar de paralelas, lo que le da a la célula un aspecto parecido al de un cono. Así pues, si estamos de acuerdo en que todo el sistema undulipoidal deriva de espiroquetas simbiontes, también lo estaremos en que la simbiosis es un requisito previo ineludible para la evolución del ojo. El patrón de selección natural por ordenador de Richard Dawkins y el estudio extensivo de la bibliografía de Ernst Mayr y sus colegas, demuestran que toda población de organismos con fotosensitividad superficial tenderá a refinar y ampliar esta capacidad en la dirección de un órgano que esté protegido, que se pueda enfocar y que incluso pueda formar una imagen. Según Mayr, los ojos han evolucionado en los linajes animales al menos cuarenta veces. Los ejemplos más aparentes y mejor estudiados son los ojos simples de los vertebrados y los moluscos (como calamares y almejas), y los compuestos de los insectos. En la relación de Mayr no estaba incluida la evolución de los ojos tipo cámara de dos géneros de protistas monocelulares (los dinomastigotes eritrodínidos), en los que la célula entera se ha transformado en un ojo funcional modificado. De hecho, el Erythrodiniopsis es completamente análogo a una lente gran angular de fotografía. Esta célula única, cuyo plástido y demás membranas recubiertas de pigmento forman el equivalente del retinal, se queda esperando a que su presa microbiana arroje su sombra sobre ella. Como en cualquier esfuerzo por buscar el origen de un rasgo complejo y superficialmente perfecto, también aquí es necesario estudiar —en palabras del propio Darwin— «las rarezas y peculiaridades» de sus antecedentes.
El lenguaje del cambio evolutivo no son ni las matemáticas ni la morfología generada por ordenador. Ciertamente, tampoco la estadística. Más bien parece que la historia natural, la ecología, la genética y el metabolismo necesitan ser suplementarios con un conocimiento fidedigno sobre los microbios. La fisiología microbiana, la ecología y la protistología son esenciales para la comprensión del proceso evolutivo. El comportamiento de los microbios, tanto dentro de sus propias poblaciones como en sus interacciones con otros, determinó el curso enrevesado y expansivo de la evolución. El mundo vivo por debajo de lo visible subyace, en última instancia, al comportamiento, el desarrollo, la ecología y la evolución del mundo de las formas de vida mucho mayores, del que formamos parte y con el que coevolucionamos. Tal vez haya quien se sienta disminuido por esta perspectiva de una evolución puntuada y movida por las uniones entre microbios pero, haciéndonos eco de las palabras de Darwin, creemos que en esta visión de la vida hay también grandeza. Aparecen formas y variaciones innumerables, pero no de forma paulatina y aleatoria, sino súbitamente y con toda su fuerza, por medio de la seducción de extraños, de la implicación y la interiorización del otro en sí mismo —vírica, bacteriana y eucariótica—, en genomas cada vez más complejos y misceláneos. La adquisición del otro reproductor, del microbio y su genoma, no es un mero espectáculo anexo. La atracción, unión, fusión, incorporación, cohabitación, recombinación —tanto permanente como cíclica— y demás acoplamientos prohibidos, son las fuentes principales de la variación que Darwin echó en falta. Sensibilidad, seducción, unión, adquisición, fusión, acomodación, perseverancia y demás capacidades de los microbios, no son en absoluto cuestiones irrelevantes para el proceso evolutivo, sino todo lo contrario. La incorporación e integración de genomas «ajenos» —bacterianos u otros— condujo a variaciones heredables significativas. La adquisición de genomas ha sido crucial en el proceso evolutivo a lo largo de la dilatada y compleja historia de la vida.
Verdaderamente, y como dijera Wallin en 1927, «no deja de resultar sorprendente la idea de que las bacterias, esos organismos que popularmente asociamos con la enfermedad, puedan representar el factor causativo fundamental en los orígenes de las especies». Estamos de acuerdo.
| Acariomastigonte | Complejo organular intracelular que se encuentra en numerosos mastigotes (células undulipoidales). El sistema mastigonte incluye el tallo axonémico y las membranas circundantes del undulipodio, el cinetosoma subyacente del que surge el undulipodio y las fibras asociadas, que pueden incluir costillas, cuerpos parabasales y axostilos. A diferencia del sistema cariomastigonte, el sistema acariomastigonte no está conectado con el núcleo vía conector nuclear o rizoplasto (figura 9.3.) |
| Amitocondriado | Un organismo eucariótico anaeróbico que carece de mitocondrias, tanto primarias (sus antepasados eucarióticos nunca adquirieron mitocondrias) como secundarias (perdió las mitocondrias que sí tenían sus antepasados). |
| Aparato de Golgi | Dictiosoma, cuerpo de Golgi. Parte del sistema endomembránico de casi todas las células eucarióticas, visible a través del microscopio electrónico, con apariencia de estructura membranosa de vesículas aplanadas o cisternas, a menudo apiladas en grupos paralelos. Implicada en la elaboración, almacenaje y secreción de productos de la síntesis celular. |
| Arqueoamebas | Una de las tres clases (arqueoamebas, metamónadas y parabasálidas) del filum arqueoprotista, las arqueoamebas incluyen amebas amitocondriadas microaerofílicas que carecen de undulipodios, así como amebas mastigo-amebasanaeróbicas que tienen undulipodios en alguna fase de su vida. |
| Arqueoprotista | Filum dentro del reino protocista constituido principalmente de amitocondriados anaeróbicos o en ocasiones protistas microaerofílicos, incluyendo amebas y mastigotes (organismos unicelulares undulipodiados) todos ellos con mitocondrias. |
| Asociación | Relación ecológica regular de dos o más clases distintas de organismos. Ver «simbiosis». |
| Bacterias | Miembros del reino bacterias (también denominado móneras o procariotas), uno de los cinco reinos de la vida. Todas las bacterias y únicamente las bacterias son procariotas, es decir, carecen de un núcleo envuelto en membrana. En ocasiones se dividen en dos subreinos: eubacterias y arqueobacterias. |
| Biólogo evolutivo | Biólogo que estudia la evolución de la vida, ya sea en general o en un organismo o unos organismos específicos, a nivel ecosistémico, ecológico, de comunidad, de población, individual, celular o molecular. El que estudia la ciencia integrada por la evolución, la ecología, el comportamiento y la sistemática. |
| Biosfera | Todos los lugares de la superficie terrestre, desde sus límites superiores en la atmósfera hasta las mayores profundidades marinas, donde existe la vida. |
| Cariomastigonte | Complejo organular intracelular distinguible en muchos mastigotes (células undulipodiadas). Orgánulos asociados con las undulipodios. El sistema mastigonte puede incluir a los cinétidos con sus undulipodios, sus membranas ondulantes, sus costillas, sus cuerpos parabasales y sus axostilos, todos ellos conectados al núcleo vía un conector nuclear, como el rizoplasto. Comparar con acariomastigonte. |
| Cariotipo | Complemento cromosómico total de un animal, una planta, un hongo o un protoctista, tal como se le puede distinguir en una preparación tintada de cromosomas condensados, a la luz del microscopio. Cariotipificar se refiere al proceso de fijar y tintar, utilizado para determinar la morfología y el número de cromosomas característicos de una especie determinada. |
| Centrómero | Estructura que liga los cromosomas a los microtúbulos del huso mitótico. Centro capturador de microtúbulos localizado en los cromosomas. Las conexiones centrómeras al huso son indispensables para la segregación cromátida. El centrómero, en cuanto región del cromosoma deducida del comportamiento genético, se distingue en ocasiones del cinetócoro como estructura observable a través del microscopio electrónico. Algunos autores consideran al centrómero como sinónimo del cinetócoro. |
| Cilio | Undulipodio. Orgánulo de motilidad que sobresale de la célula, formado por un axonema cubierto por la membrana del plasma. El término se utiliza para referirse a los undulipodios de los ciliados y de las células de los tejidos de los animales. Compuesto de [9(2)+2] tallos micro tubulares o axonemas. |
| Cinetócoro | Centro organizador de microtúbulos, localizado habitualmente en una sección hundida del cromosoma, que mantiene unidos a los cromátidos. Los cinetócoros, manifestaciones morfológicamente visibles de los centrómeros, constituyen el punto de anclaje de las fibras que forman el huso durante la división del núcleo (mitosis y meiosis). Los centrómeros se deducen del comportamiento genético, mientras que los cinetócoros son directamente visibles por medio del microscopio electrónico. |
| Cinetosoma | Orgánulo intracelular, no delimitado por membrana alguna, característico de todas las células undulipodiadas. Estructuras micro tubulares necesarias para la formación de undulipodios. Los cinetosomas difieren de los centríolos en que a partir de ellos se extiende la tija o axonema. Sus microtúbulos están organizados según el patrón [9(3)+0]. Todos los undulipodios están sustentados por cinetosomas. Estos orgánulos básales, frecuentemente denominados «cuerpos básales» porque generan el axonema, son necesarios para la formación de todos los undulipodios. Los cinetosomas difieren de los centríolos en que a partir de ellos se extienden axonemas tipo [9(2)+2]. Debido a su mayor precisión, el término «cinetosoma» es preferible al de «cuerpo basal». |
| Clasificación | Proceso de establecimiento, definición y clasificación de taxones en series jerárquicas de grupos, tanto artificiales como naturales. Un esquema de estas series de grupos o taxones. |
| Conector nuclear (rizoplasto) | Banda microfibrilar cruzada, que se extiende desde las bases de los cinetosomas y se dirige hacia el núcleo o hacia los centros citoplásmicos organizadores de microtúbulos. Sistema celular organular. |
| Consorcio | Grupo de individuos de especies distintas, típicamente de diferentes filia, que viven en asociación estrecha. En microbiología, asociación física entre las células de dos o más tipos de microorganismos distintos, más o menos estable y ventajosa, como mínimo para una de las partes. |
| Crecimiento exponencial | Crecimiento sin obstáculos, de modo que cuanto mayor es la población más rápidamente crece ésta. Incremento geométrico de una población que creciera en un entorno ideal, sin limitaciones. |
| Epibionte | Término ecológico que describe la topología de la asociación entre organismos, en la que uno de ellos (el epibionte) vive sobre la superficie de otro. |
| Epífito | Planta que crece en otra planta (forófito) por razones de apoyo o anclaje, más que para conseguir agua o nutrientes. Cualquier organismo que viva sobre la superficie de una planta. |
| Escala temporal geológica | Escala temporal, establecida por los geólogos, que refleja los cambios en la historia geológica del planeta. Los eones constituyen sus divisiones mayores: Hadeano, Arqueozoico, Proterozoico y Fanerozoico. La paleontología tradicional tiende a centrar su atención únicamente en los acontecimientos macroscópicos del Fanerozoico (ver tabla 9.1) |
| Especie | Grupo de organismos, minerales u otras entidades, formalmente reconocido como distinto de otros grupos. Uno de los taxones del catálogo de especies. En la jerarquía de la clasificación biológica, la categoría que ocupa el lugar inmediatamente inferior al género. Unidad básica de clasificación biológica. En este libro sugerimos que son miembros de una misma especie los organismos con la misma clase y el mismo número de genomas integrados en común. |
| Eucariotas | Organismos que tienen células nucleadas, células con al menos un núcleo envuelto en membrana que sigue algún tipo de mitosis. Todos los eucariotas (reinos protoctista, animalia, hongos y plantas) derivan de, e incluyen a, los protoctistas. El reino bacterias (procariotas) carece de células nucleadas. |
| Euplotidium | Protista ciliado (con bacterias proyectables). |
| Euprymna scolopes | Denominado comúnmente calamar hawaiano rabicorto. Alberga la bacteria luminiscente simbionte Vibrio fischeri. La evolución simbiótica ha conducido a la emergencia del complejo órgano emisor de luz del calamar, en el que están contenidas las bacterias. |
| Evolución | Cualquier cambio acumulativo en las características de organismos o poblaciones, de generación en generación. Descendencia o desarrollo con modificaciones. Todos los cambios que han transformado la vida sobre la Tierra, desde sus inicios hasta la diversidad que la caracteriza en el presente. |
| Filogenia | Secuencia hipotética de ascendencia de grupos de organismos, tal como la reflejan sus respectivos árboles evolutivos. Comparar con «sistemática» y «taxonomía»—. |
| Flagelo | Flagelo bacteriano: estructura procariótica extracelular, compuesta de polímeros proteicos homogéneos, pertenecientes a la clase de proteínas denominadas flagelinas. Se mueve por rotación sobre su base. Tija relativamente rígida impulsada por un motor rotatorio embebido en la membrana celular, intrínsecamente no mótil y ocasionalmente enfundado. Término utilizado a veces erróneamente para referirse al undulipodio eucariótico, o estructura intracelular intrínsecamente motil, utilizada por los eucariotas para desplazarse y alimentarse, compuesta por una disposición estándar de nueve microtúbulos dobles y dos microtúbulos centrales compuestos por tubulina, dineína y aproximadamente otras 200 proteínas, entre las que no se encuentra la flagelina. Ningún flagelo (pero sí todos los undulipodios) es sustentado por un cinetosoma. |
| Fotoautotrofia | Modalidad de nutrición en la que la luz constituye la fuente de la energía. Un organismo obligadamente fotoautotrófico utiliza la energía de la luz para sintetizar material celular a partir de compuestos inorgánicos (dióxido de carbono, sales de nitrógeno). |
| Fotosíntesis | Proceso por el que se genera energía química a partir de la luz solar. En él se forman determinados carbohidratos hexosos a partir del dióxido de carbono y del agua en los citoplastos de las células de los vegetales vivos, con oxígeno o azufre como residuos resultantes de los compuestos agua y sulfuro de hidrógeno, que aportan el hidrógeno. |
| Gérmenes | Término no científico, utilizado para referirse a microbios que pueden o no causar enfermedades. Término empleado para referirse a los microbios como algo sucio, que la gente quiere evitar. «Enemigos» asociados a las enfermedades contagiosas. No distingue entre virus, bacterias, protistas, priones, etc. |
| Gunnera | Único grupo de angiospermas (plantas con flores) que forma asociaciones simbióticas regulares con cianobacterias. Existen cuarenta y tres especies dentro del género Gunnera, todas ellas asociadas con la bacteria fijadora de nitrógeno Nostoc. La especie mayor o G. Manicata, se conoce popularmente con el nombre de «paraguas del pobre». |
| Hermissenda | Género de moluscos nudibranquios (babosas marinas) que presentan asociación simbiótica con cnidarios tubiformes, animales marinos sésiles que poseen células urticantes denominadas nematocistos. (Medusas, corales y anémonas de mar pertenecen también al filum Coelenteratá). Los nudibranquios, que pueden alimentarse de los hidroides tubulares sin ser picados por sus nematocistos, almacenan estas células urticantes en órganos especializados, como medio de defensa. |
| Heterotermes tenuis | Termita subterránea comedora de madera, perteneciente a la familia rhinotermitidae. Muy extendida en ambas Américas. Se sabe que alberga en su intestino protistas simbiontes, tales como el Holomastigotoides, Pseudotrichonympha y Spirotrichonympha . |
| Híbrido | Descendiente de un cruce entre individuos genéticamente distintos. Restringido a menudo en taxonomía a la descendencia de cruces interespecíficos. Una comunidad que incluye taxones derivados de dos o más comunidades distintas. En el metabolismo de ácidos nucleicos, un polinucleótido de doble hebra, en el que una hebra es ADN y la otra ARN. |
| Larva | Forma autónoma, sexualmente inmadura, en el ciclo vital de algún animal, que puede diferir de la forma adulta en morfología, nutrición y hábitat. |
| Mendel, Gregor Johann | Monje austríaco (1822-1884), padre de la genética de Mendel. Sus principios estáticos de genética, el de dominancia en un heterocigoto (un alelo puede ocultar la presencia de otro) y el de segregación en un mismo heterocigoto (dos alelos diferentes se segregan durante la formación de los gametos), combinados con la idea darwiniana del cambio a través del tiempo, formaron la visión de la evolución (neodarwinista) característica de la genética de poblaciones. |
| Metabolismo | La suma de procesos químicos y físicos que ocurren en los organismos vivos e implican una continua sustitución de sus constituyentes químicos. |
| Metamónada | Una de las tres clases (arqueoamebas, metamónadas y parabasálidas) del filum arqueoprotista. Las metamónadas son protistas amitocondriados anaeróbicos con cariomastigontes. Sus núcleos están ligados a sus undulipodios mediante conectores nucleares. La Giardia es una metamónada independiente, causante de enfermedades. Muchas viven simbióticamente en el intestino de los insectos. |
| Microbios (Microorganismos) | Seres (habitualmente bacterias, protoctistas y hongos) que se ven mejor mediante el microscopio. |
| Modalidad metabólica | La clase de metabolismo que tiene un organismo, basándose en si utiliza la luz o los compuestos químicos para obtener energía, tanto si, como fuente de electrones, utiliza compuestos orgánicos como inorgánicos, tanto si utiliza el CO2como si ingiere alimentos como fuente de carbono. |
| Moluscos | Filum muy diverso de animales, formado por organismos de agua salada, de agua dulce y terrestres. La mayoría de moluscos tiene un cascarón interno o externo, un pie muscular y un cuerpo segmentado y blando. Incluye a las almejas, a los nautilos, a los calamares, a los pulpos, a los caracoles, a las babosas y a los nudibranquios. |
| Monofilia | Condición o rasgo de un grupo de organismos, considerados como evolucionados a partir de un único antepasado común. Se dice de los taxones hermanos que son monofiléticos. |
| Monoteísmo | Creencia religiosa en un dios único, sobrenatural y todopoderoso. El judaísmo, el cristianismo y el islam constituyen ejemplos de religiones monoteístas. |
| Nematocisto | Cnidocisto. Célula modificada con una cápsula que contiene un aguijón filamentoso utilizado como elemento de defensa, anclaje o captura. Algunos contienen sustancias venenosas o paralizantes (por ejemplo, en todos los celentéreos y ctenóforos). Se encuentran orgánulos análogos en algunos dinomastigotes y ciliados cariorrelictos y suctorios. |
| Neodarwinismo | Creencia en que la mutación aleatoria constituye la fuente principal de cambio evolutivo, sobre la que actúa la selección natural. Deriva de la fusión entre la genética de Mendel y la “descendencia con modificación” de Darwin. |
| Núcleo | Orgánulo esférico, envuelto por una membrana, que contiene ADN. Universal en protoctistas, animales, plantas y hongos. Cromatina (ADN y proteína) organizada en cromosomas. Lugar donde ocurre la síntesis del ADN y la transcripción del ARN. Órgano definitorio de los eucariotas. |
| Nucleoide | Estructura que contiene ADN en los procariotas, no circunscrita por la membrana celular. No contiene poros nucleares. |
| Nudibranquio | Molusco gasterópodo sin cáscara, como la babosa marina. (Los buccinos y los caracoles terrestres son también moluscos gasterópodos). |
| Nueva síntesis (Síntesis moderna) | Teoría comprensiva de la evolución, que enfatiza la selección natural, el gradualismo y las poblaciones como unidades fundamentales para el cambio evolutivo. Neodarwinismo. |
| Olenus , olénidos | Taxón de trilobites del Paleozoico Inferior que vivió en los fondos marinos, bajo condiciones de escasez de oxígeno y abundancia de azufre. Abundante en la región donde actualmente está Escandinavia. Estas áreas no sustentaron ninguna otra forma de vida macroscópica. Se cree que los trilobites olénidos pudieron vivir en este medio difícil gracias a bacterias quimioautotróficas simbiontes. |
| Parabasálidas | Una de las tres clases (arqueoamebas, metamónadas y parabasálidas) del filum arqueoprotista. Las parabasálidas son protistas simbiontes que viven en los intestinos de los insectos. Aparentemente digieren la celulosa, a partir de la cual extraen azúcares. Cada parabasálida tiene al menos cuatro undulipodios, un axostilo y visibles cuerpos básales. Los dos órdenes de parabasálidas son tricomonádidas e hipermastígidas. |
| Potencial biótico | El número de organismos que pueden ser producidos en una única generación, o unidad de tiempo, característico de determinada especie, medido en el número máximo de descendientes por generación, número máximo de esporas producidas por año o términos equivalentes. Ilustra las tendencias de los organismos a aumentar exponencialmente cuando se dan las condiciones adecuadas para el crecimiento material. |
| Procariotas | Bacterias. Miembros del reino de las Bacterias (reino mónera o procariota). Organismo celular compuesto de células con nucleoides, con ausencia de núcleo circunscrito por membrana (ver ejemplo en figura 9.2). |
| Protista | Cada célula de los miembros con pocas células, y por consiguiente microscópicos, del reino protoctista. |
| Protoctista | Uno de los cinco reinos en los que se clasifican todos los organismos vivos. Los protoctistas son los organismos eucarióticos nucleados (la célula única protista), así como sus descendientes directos multicelulares. Este reino incluye a todos los organismos eucarióticos, a excepción de animales, plantas y hongos, por ejemplo, todas las algas, los mohos del lodo, las amebas, las redecillas del lodo, los mohos acuáticos y las foraminíferas se encuentran entre las 250.000 especies existentes en unos cincuenta grupos principales (ver ejemplos en figuras 7.4, 7.5 ,9.1,9.3 y 9.4). |
| Quimera | Un organismo que reúne dos o más tipos genéticos, formado por simbiosis, segregación anormal de cromosomas o implante artificial. Término utilizado habitualmente en referencia a microorganismos y plantas, raramente a animales. Un mosaico. |
| Quimioautotrofia | Modo metabólico de nutrición por el que un organismo obtiene energía por medio de la oxidación de substratos inorgánicos tales como el azufre, el nitrógeno o el hierro, junto con carbono celular a partir de dióxido de carbono (CO2). |
| Selección natural | Proceso resultante de la producción de más organismos de los que pueden sobrevivir. Se dice que los que sobreviven y tienen descendencia han sido seleccionados naturalmente. |
| Simbiosis | Asociación física prolongada entre dos o más organismos «distintamente denominados», generalmente de dos o más especies diferentes. Los niveles de integración asociativa en la simbiosis pueden presentar diversos grados de intimidad. La integración puede ser por metabolismo, por productos genéticos o genética. Los miembros de una misma simbiosis son simbiontes entre sí. |
| Sistemática | Subcampo de la ciencia evolutiva que se ocupa de la denominación, clasificación y agrupación de organismos, en base a sus relaciones evolutivas. Comparar con «filogenia» y «taxonomía». |
| Snyderella | Protista multinucleado, simbionte de la termita. Calonínfido (figuras 7.4 y 9.4. |
| Staurojoenina | Protista hipermastigote simbionte de la termita (ver figura 7.5.) |
| Taxón | Grupo de organismos similares, como reino (taxón más inclusivo), filum, familia, género o especie (taxón menos inclusivo). |
| Taxonomía | Teoría y práctica de descripción, denominación y clasificación de organismos. Comparar con «sistemática» y «filogenia». |
| Thiodendron | Espiroqueta de Dubinina en asociación con una bacteria reductora del azufre. Consorcio marino originalmente descrito como una sola bacteria. |
| TVilobites | Grupo extinto de animales que vivieron durante el Cámbrico. Artrópodo segmentado, de cáscara dura. Fósil común. |
| Undulipodio | Orgánulo de motilidad cubierto de membrana celular, que exhibe en ocasiones funciones sensoriales o de alimentación, compuesto por al menos 200 proteínas. Axonema microtubular [9(2)+2], habitualmente cubierto con membranas de plasma, circunscrito a las células eucarióticas. Incluye cilios y colas de espermatozoides. Cada undulipodio se desarrolla invariablemente a partir de su cinetosoma, estructura microtubular simétrica de nueve partes en la base. Contrasta en numerosos aspectos con el orgánulo de motilidad procariótico o flagelo, estructura rígida compuesta por una única proteína. La bibliografía biológica se refiere frecuentemente al undulipodio con el término desfasado de “flagelo” o “euflagelo”. Sinónimo: flagelo eucariótico. |
| Zoocentrismo | Estudio de los animales, incluyendo los humanos, como si los animales fueran los principales organismos existentes, así como y/o los únicos merecedores de estudio y de financiación para su investigación. Gran displicencia para con los otros cuatro reinos de la vida, considerándolos ignorantemente como propios de formas de vida «menores». Ignora las influencias fundamentales que los miembros de esos otros reinos ejercen sobre el reino animal, el medio natural y los ecosistemas planetarios. |
- Arsatt, Peter, 2002. Hipótesis micosómica: los hongos se propagan dentro de plástidos de tejido de plantas senescentes. (Manuscrito sin publicar).
- Behe, Michael, 1996. Darwin’s Black Box. Free Press, Nueva York. [Versión en castellano: La caja negra de Darwin: el reto de la bioquímica a la evolución. Barcelona: Andrés Bello, 2000.]
- Brodo, I. M., S. D. Shamoff y S. Shamoff, 2001. Lichens of North America. Yale University Press, New Haven.
- Caldwell, D. E., 1999. Post-modern ecology: Is the environment the organism? Environmental Microbiology 1:279-281.
- Chapman, M. J., M. F. Dollan y L. Margulis, 2000. Centrioles and kinetosomes: Form, function and evolution. Quarterly Review of Biology 75:409-429.
- Diamond, Jared, 1999. Guns, germs and Steel: the fates of human societies. W. W. Norton, Nueva York. [Versión en castellano: Armas, gérmenes y acero. Madrid: Debate, 1998.]
- Dolan, M., 2001. Speciacion of termite gut protists: The role of bacterial symbionts. International Microbiology 4:203-218.
- Dolan, M., H. Melnitsky, R. Kolnicki y L. Margulis, 2002. Motility proteins and the origin of the nucleus. Anatomical Record (en imprenta).
- Fortey, Richard, 1998. Life: An unauthorized biography. Oxford University Press, Oxford, Reino Unido. [Versión en castellano: La vida: una biografía no autorizada. Madrid: Taurus, 1999.]
- Fortey, Richard, 2000. Olenid trilobites: The oldest known chemoautotrophic simbyonts? Proceedings of the National Academy of Sciences 97:6574-6578.
- Fortey, Richard, 2000. Trilobite! Eyewitness to evolution. Flamingo, con impresión de HarperCollins, Londres.
- Freeman, S. y J. C. Herrón, 2001. Evolutionary analysis. Segunda edición. Prentice-Hall, Englewood Cliffs, N. J. [Versión en castellano: Análisis evolutivo. Madrid: Pearson Educación, 2002.]
- Harold, Franklin, 2001. The way of the cell. Oxford University Press, Nueva York.
- Hoffman, Paul F. y Daniel P. Schrag. Snowball Earth. Scientific American, enero del 2000:68-75.
- Keller, E. F. y E. A. Lloyd, 1992. Key words in evolutionary biology. Harvard University Press, Cambridge.
- Keller, Laurent, 1999. Levels of selection in evolution. Princeton University Press, Princeton.
- Khakhina, L. N., 1992. Concepts of symbiogenesis: A historial and critical study of the research of Russian botanists . Yale University Press, New Haven.
- King, Jennifer M., Tom S. Hays y R. Bruce Nicklas, 2000. Dyenin is a transient kinetochore component whose binding is regulated by microtubule attachement, not tensión. Journal of Cell Biology 151:739-748.
- Kolnichi, Robin, 1999. Karyotipic fission theory applied: Kinetochore reproductions and lémur evolution. Symbiosis 26.123-141.
- Kolnichi, Robin, 2000. Kinetochore reproduction in animal evolution: Cell biological explanation of karyotipic fission theory. Proceedings of the National Academy of Sciences 97:9493-9497.
- Kolnichi, Robin, Kinetochore reproduction (=karyotipic fission) theory: Divergence of lémur taxa subsequent to simple chromosomal mutation. Evolution (en preparación).
- Lapo, Andre, 1987. Traces of bygone biospheres. Mir, Moscú.
- Lovelock, J. E., 1988. The Ages of Gaia: A biography on our living Earth. W. W. Norton, Nueva York. [Versión en castellano: Las edades de Gaia. Barcelona: Tusquets, 1993.]
- Lovelock, J. E., 2000. Homage to Gaia: The life of an independent scientist. Oxford University Press, Oxford, Reino Unido.
- Marcotrigiano, Michael, 1999. Chimeras and variegation: Pattems of deceit. Horticultural Science 32:773-784.
- Margulis, Lynn, 1998. Symbiotic planet. A new look at evolution
- Basic Books, Nueva York. [Versión en castellano: Planeta simbiótico. Un nuevo punto de vista sobre la evolución. Madrid: Debate, 2002.]
- Margulis, Lynn, 1990. Words as battlecries: symbiogenesis and the new field of endocytobiology. BioScience 40:673-677.
- Margulis, L., M. F. Dolan y R. Guerrero, 2000. The chimeric eukaryote: Origin of the nucleus from the karyomastigonts in amitochondriate protists. Proceedings of the National Academy of Sciences 97:6954-6959.
- Margulis, L. y M. F. Dolan, 2002. Early Ufe. Segunda edición. Jones and Bartlett, Sudbury, Massachusetts.
- Margulis, L. y L. Olendzenski, 2000. Microcosmos, Vol. 1 (Células y Reproducción) y Vol. 2 (Evolución y Diversidad). Vídeos y sus folletos correspondientes. Jones and Bartlett Publishers, Boston.
- Margulis, L. y Dorion Sagan, 1996. Gaia y microcosmos. Vol. 1. A Sciencewriters Vídeo.
- Margulis, L. y Dorion Sagan, 1997. Microcosmos: Four billion years of microbial evolution. University of California Press, Berkeley. [Versión en castellano: Microcosmos . Barcelona: Tusquets, 1995.]
- Margulis, L. y Dorion Sagan, 1999. Slanted truths: Essays on Gaia, symbiosis and evolution. Copemicus Springer-Verlag, Nueva York. [Versión parcial en castellano: Una revolución en la evolución. Valencia: Universidad de Valencia, 2003.]
- Margulis, L. y Dorion Sagan, 2000. What is life? University of California Press, Berkeley. [Versión en castellano: ¿Qué es la vida? Barcelona: Tusquets, 1996.]
- Margulis, L. y K. V. Schwartz, 1998. Five kingdoms: An illustrated guide to the phyla of life on Earth. Tercera edición. W. H. Freeman and Company. [Versión en castellano: Cinco reinos. Barcelona: Labor, 1985.]
- Mayr, E., 1942. Systematics and the origin of species. Harvard University Press, Cambridge, Massachusetts.
- Mayr, E., 1982. Growth of biological thought. Harvard University Press, Cambridge, Massachusetts.
- Mayr, E., 2001. What evolution is. Basic Books, Nueva York.
- Mayr, E. y J. Diamond, 2001. The birds of northern Melanesia: speciation, ecology and biogeography . Oxford University Press, Oxford, Reino Unido y Nueva York.
- Mekos, D. C., 1992. Apéndice: Ivan E. Wallin and his theory of symbionticism. En Khakhina, L. N., Concepts of symbiogenesis: A historical and critical study of Russian botanists . Yale University Press, New Haven.
- Miklos, G. L. G., 1993. Emergence of organizational complexities during metazoan evolution: perpectives from molecular biology, paleontology and neo-Darwinism. - - - Membership of Australian Paleontologists, N o. 15, págs. 7-14.
- Morrison, Reg, 1999. The spirit in the gene: Humanity’s proud delusion and the laws of nature . Cornell University Press, Nueva York.
- Nicklas, Bruce, M. S. Campbell, S. C. Ward y G. J. Gorbsky, 1998. Tension-sensitive kinetochore phosphorylation in vitro. Journal of Cell Science. 111:3189-3196.
- Oheler, Stefan y Kostas Bourtzis, 2000. Primera conferencia internacional Wolbachia: Wolbachia 2000. Symbiosis 29:151-161.
- Olendzenski, Lorraine, Lynn Margulis y Steven Goodwin, 1998. Looking at microbes: A microbiology laboratory for students. Jones and Bartlett Publishers, Boston.
- Patouillard, N. y A. Gaillard, 1888. Champignons du Vénézuéla et principalment de la région du Haut Orénoque récoltés en 1887 par M. A. Gaillard. Bulletin de la Societé Mycologique de France 4:7-46.
- Polz, M. F., J. A. Ott, M. Bright y C. M. Cavanaugh, 2000. When bacteria hitch a ride. ASM news 66:531-538.
- Rumpho, M. E., E. J. Summer y J. R. Meinhart, 2000. Solar-powered sea slugs. Mollusc/algal chloroplast symbiosis. Plant Physiology 123:29-38.
- Sagan, D. y E. D. Schneider, 2000. The pleasures of change en The forres of change. National Geographic Society, págs. 115-126.Washington, D. C.
- Sapp, J., 1994. Evolution by association: A history of symbiosis. Oxford University Press, Nueva York.
- Sapp, J., 2000. Génesis: The evolution of biology. Oxford University Press, Nueva York.
- Schaechter, M., 1997. In the company of mushrooms. Harvard University Press, Cambridge, Massachusetts. [Versión en castellano: En compañía de las setas. Barcelona: Reverté, 1999.]
- Schneider, E. D. y J. J. Kay, 1994. Life as a manifestation of the second law of thermodynamics. Mathematical Computer Modeling 19, N o. 6-8:25-48.
- Sciencewriters, 2002. Página electrónica: Animales verdes y otros vídeos sobre simbiosis a consultar en esta página.
- Shapiro, James A., 1998. Bacteria as multicellular organisms. Scientific American 256:82-89.
- Shostak, S. y V. Kolluri, 1995. Symbiotic origins of cnidarian cnidocysts. Symbiosis 19:1-19.
- Smith, D. C. y S. E. Douglas, 1987. The biology of symbiosis. E. Arnold, Londres.
- Sonea, S. y L. Mathieu, 2000. Prokaryotology. Les Presses de l’Université de Montreal, Montreal.
- Todd, Neil B., 1970. Karyotipic fissioning and canid phylogeny. Journal of Theoretical Biology 26; 445-480.
- Todd, Neil B., 2000. Mammalian evolution: Karyotipic Fission Theory. En L. Margulis, C. Mathews y A. Haselton, eds., Environmental Evolution. Segunda edición. MIT Press, Cambridge, Massachusetts y Londres.
- Todd, Neil B., 2001. Kinetochore reproduction underlies karyotypic fission theory: Possible legacy of symbiogenesis in mammalian chromosome evolution. Symbiosis 29: 319-327.
- Tumer, J. Scott, 2000. The extended organism: The physiology of animal-built structures. Harvard University Press, Cambridge, Massachusetts.
- Vemadsky, V. I., 1998. The biosphere (en inglés), traducción del original en ruso de Vemadsky La biosfera, 1926). Copemicus Springer-Verlag, Nueva York. [Versión en castellano: La biosfera. Madrid: Fundación Argentaria/Visor, 1997.]
- Wallin, I. E., 1927. Symbionticism and the origin of species. Williams and Wilkins, Baltimore.
- White, M. J.D., 1973. Animal cytology and evolution. Cambridge University Press, Cambridge, Reino Unido. [Versión en castellano: Citología animal y evolución. Buenos Aires: Espasa Calpe, 1951.]
- Wiener, Jonathan, 1999. The beak of the finch. Harvard University Press, Cambridge, Massachusetts.
- Williamson, D. I., 1992. Larvae and evolution: toward a new zoology. Chapman y Hall, Nueva York.
- Williamson, D. I., 2001. Larval transfer and the origin of larvae. Zoological Journal of the Linnean Society 131:111-122.
- Wright, Barbara E., 2000. A biochemical mechanism for nonrandom mutations and evolution. Journal of Bacteriology 192:2993-3001.
Entre los colegas de profesión que nos han ayudado con el manuscrito, tanto de forma directa como indirecta, cabe cita a Marcus Aebi, Jennifer Benson, Peter G. Brown, Lois Brynes, Emily Case, Michael Chapman, Michael Dolan, Ann Ferguson, June Girard, Brianne Goodspeed, Jennifer Gotlieb, Ricardo Guerrero, Jessie Gunnard, Kenneth Hsu, Michael Keston, Alan Kuzerian, Tom Kunz, Wolfgang Krumbein, James Lovelock, Adam McConnell, Kay Mariea, Hannah Melnitsky, Vanessa Mobley, Carlos Montufar, Marta Norman, Morriss Partee, Simon Powell, Donna Reppard, Jan Sapp, Eric D. Schneider, Judith Serrin, Dean Soulia, Crispin Tickell, Gabriel Trueba, Peter Warshall, Andrew Wier, y Sean Werle. Los trabajos artísticos de Christie Lyons y Kathryn Delisle, entre otros, adornan estas páginas, junto con fotografías de artistas como Reg Morrison. Tenemos una deuda particular para con Judith Herrick (Typro), que mecanografió originales y revisiones con gran presteza, precisión y alegría. Le agradecemos igualmente a Bill Frucht sus penetrantes preguntas y sus esclarecedores comentarios, que contribuyeron a modelar y editar toda la obra, desde su etapa como propuesta hasta su fase final como libro terminado, así como a Ernst Mayr por su crítica superior en la hora veinticinco. Las ayudas financieras necesarias, tanto para nuestros estudios como para nuestras publicaciones, procedieron de la NASA Space Sciences, la Richard Lounsbery y de la Universidad de Massachusetts Graduate School.
La hospitalidad del Collegium Helveticum (Zurich) y del Hanse Wissenschaft Kolleg (Dalmenhorst, Alemania) fueron esenciales para la terminación de esta obra, como lo fue la ayuda de la Humboldt Foundation, hacia la que nos sentimos particularmente agradecidos.
Notas: