
De las celulas a las civilizaciones
Enrico Coen
Introducción:
Una receta para los cambios
§. Historia y formaLa vida tiene un poder transformante muy notable: a lo largo de miles de millones de años de evolución, las formas elementales se han transformado en las criaturas complejas de hoy en día. Durante nueve meses, un cigoto indefinido se convierte en un ser humano. Durante unos pocos años, un bebé de movimientos imprecisos se convierte en un adulto que camina, habla y razona. Y durante diez mil años, las sociedades humanas han pasado de las pequeñas comunidades tribales a las ciudades y civilizaciones complejas de hoy en día.
§. Receta creativa para la vida
Resulta tentador pensar que tras esta propiedad transformadora de la vida existe un solo mecanismo, pero cuando la estudiamos vemos en funcionamiento cuatro mecanismos muy diferentes. Se cree que todas las criaturas de nuestro planeta tuvieron su origen en la competencia entre individuos por la supervivencia y la reproducción durante muchas generaciones, mediante la selección darwiniana. En el desarrollo de un cigoto interviene un mecanismo realmente diferente ya que, una vez fecundado, pasa por varios ciclos de división en los que se van formando patrones en el embrión mediante señales celulares y las diferentes maneras de responderlos. El desarrollo tiene que ver más con la formación de patrones dentro de un embrión en crecimiento que con la competencia por el éxito reproductor. El aprendizaje depende de otro mecanismo: a medida que un animal interacciona con su entorno, las conexiones neuronales se modifican en el cerebro. Algunas conexiones se pierden o se debilitan, mientras que otras se forman o se refuerzan, lo que permite la captura de nuevas relaciones con el entorno, de manera que el aprendizaje no es más que la modificación de las interacciones y conexiones neurales. Finalmente llegamos al cambio cultural: los humanos interaccionan dentro de los grupos sociales, lo que conduce a avances relacionados con herramientas, utensilios y conocimiento. La cultura es un fenómeno social que depende de la forma en la que nos comportamos y en la que interaccionamos con los demás.
No existe ningún punto en común obvio entre el funcionamiento de estos diferentes procesos, pues todos parecen transcurrir por caminos completamente diferentes: la evolución mediante diferencias en el éxito reproductor; el desarrollo mediante la proliferación, el crecimiento y formación de patrones celulares; el aprendizaje mediante cambios en las conexiones neuronales; y el cambio cultural a través de las interacciones humanas.
Parece extraño que en la naturaleza hayan adquirido importancia cuatro mecanismos de transformación totalmente diferentes. Así como los físicos se afanan por elaborar una «teoría de todo» que reúna sus teorías fundamentales, se espera que los biólogos busquen una teoría unificada para las transformaciones que afectan a los seres vivos, una teoría que englobe evolución, desarrollo, aprendizaje y cambio cultural. En el pasado ya se había intentado dicha unificación: Ernest Haeckel, un seguidor entusiasta de Darwin en el siglo XIX, propuso una relación directa entre la evolución y el desarrollo. Pensaba que a medida que se desarrolla un embrión, se recrea su historia evolutiva. Así pues, los embriones humanos pasarían por una etapa pisciforme, otra reptiliana, y así sucesivamente, a medida que crecen en el vientre. Sin embargo, más tarde se demostró que esta idea estaba desencaminada[1]. Más recientemente, Gerald Edelman ha intentado integrar la evolución y el aprendizaje en su teoría del «darwinismo neural»[2]. En la década de los ochenta del siglo XX, Edelman propuso que las estructuras neuronales del cerebro se seleccionan durante el aprendizaje, en claro paralelismo con la selección natural, aunque recibió críticas demoledoras. Parece que la explicación unificada de diferentes transformaciones que afectan a los seres vivos está plagada de dificultades.
Quizá la naturaleza tenga realmente cuatro maneras completamente diferentes de transformarse a sí misma, sin que debamos preocuparnos más, pero creo que se trata de una perspectiva errónea. En este libro quiero mostrar que los últimos avances de nuestro conocimiento científico nos han dado acceso a una imagen unificada de cómo los sistemas vivientes se transforman a sí mismos, desde el origen de las bacterias hasta la creación de una obra maestra artística. Por primera vez comenzamos a ver el conjunto común de ingredientes y mecanismos que son responsables de las transformaciones que afectan a los seres vivos.
¿Por qué debe importarnos el hallazgo de los ingredientes comunes? Después de todo, los estudios sobre la evolución, el desarrollo, el aprendizaje y la cultura hasta la fecha parecen haber progresado muy alegremente sin preocuparse por las similitudes que comparten. ¿Qué ganamos con verlos de forma colectiva? Supongamos que comparamos el modo en que el hielo se funde con la manera en que el agua hierve. Ambos procesos difieren en muchos aspectos: el primero se refiere a un sólido que se vuelve líquido en torno a los 0 °C, mientras que el segundo afecta a un líquido que se convierte en gas a 100 °C. Se puede ver que ambas transiciones tienen muchas características en común, puesto que en ambas interviene un cambio de la fuerza y de la energía de las interacciones entre las moléculas de agua. Se trata de manifestaciones diferentes del mismo proceso subyacente. Esta perspectiva unificadora nos ayuda a comprender mejor lo que está ocurriendo que si nos limitáramos a estudiar simplemente cada transición por separado. Del mismo modo, el contemplar los elementos comunes que subyacen en las diferentes transformaciones de lo viviente nos ayuda a conocer la esencia de cada proceso, a la vez que nos amplía la visión de conjunto de los sucesos.
Esta estrategia quizá sea razonable para la evolución, para el desarrollo y para el aprendizaje al ser todos ellos objeto de continuas investigaciones científicas, pero no parece probable que deba extenderse al cambio cultural. Tendemos a pensar que la creatividad y la cultura humanas nos resultan tan complicadas y especiales que la ciencia tiene poco que decir sobre ellas. Pero cuando nos ponemos a ver las transformaciones de lo viviente en conjunto, comprobamos que la ciencia desempeña dos funciones. Por una parte, la ciencia proporciona una fuente de conocimiento sobre el mundo y nuestro lugar en él, y así enmarca nuestra cultura. Por otra parte, la ciencia es un producto de la cultura, el resultado de la colaboración entre los humanos durante muchos años para dar sentido al mundo que nos rodea. Solo cuando estudiemos a la vez todos los tipos de transformaciones de lo viviente seremos capaces de obtener una perspectiva clara de este doble aspecto de nuestro punto de vista, de cómo la ciencia enmarca a nuestra cultura y es enmarcada por ella. Entonces no solo comprenderemos mejor cómo se produjo el cambio cultural, sino también cómo se relaciona con nuestro pasado biológico.
¿Por qué ha costado tanto llegar a este punto de vista colectivo?
§. Historia y forma
A primera vista, la guerra y el ajedrez son muy diferentes: en la guerra hay personas que luchan y se matan entre sí, mientras que en el ajedrez hay dos personas sentadas pacíficamente frente a un tablero y que empujan algunas piezas de madera por él. Aun así, a pesar de tan obvias diferencias, las dos están muy relacionadas. Primero, están conectadas en la historia: el origen del ajedrez se puede rastrear hasta el juego del shatranj jugado en Persia entre los siglos V y VI, que a su vez podría proceder del juego indio chaturanga[3]. Al igual que el ajedrez moderno, el shatranj era un juego para dos jugadores con 32 piezas en un tablero de 64 escaques (casillas). Cada jugador tenía un ejército compuesto por dos elefantes, dos caballos, dos carros de combate y ocho soldados a pie. Se basaban en las principales unidades de lucha de la época y eran los predecesores de los alfiles, caballos, torres y peones del ajedrez moderno. También había un rey y un ministro (equivalente a la reina moderna). El objetivo era capturar o atrapar al rey del oponente. El juego ilustraba cómo un ejército podría vencer y flanquear a otro a base de estrategia y sagacidad. En su época, el shatranj era tan juego de guerra como hoy en día lo son los de ordenador.Hay otras maneras de relacionar el ajedrez con la guerra que no dependen del conocimiento de sus conexiones históricas. Ambos se pueden considerar muy competitivos, donde un adversario intenta derrotar a otro. Ambos son territoriales, y cada adversario intenta ocupar o controlar regiones. En ambos se intenta eliminar o capturar los elementos de la oposición. Y hay una fuerte componente estratégica de cooperación lateral, con unidades que se apoyan unas a otras cuando montan un ataque o mantienen una defensa. La guerra y el ajedrez tienen formas parecidas además de estar conectados por la historia.
Estos dos modos de relacionar el ajedrez con la guerra (por la historia o por la forma) están a su vez interrelacionados. Las similitudes de forma, como la competición y la territorialidad, se deben a los orígenes del ajedrez como juego que simula la guerra. Sin embargo, en el ajedrez no aparecen todas las peculiaridades de la guerra: no aparecen elementos como la muerte física de humanos, ni se varía el trazado del terreno ni las condiciones ambientales. El ajedrez siempre se juega con la misma distribución de escaques y las condiciones iniciales están perfectamente controladas, sin que participe en el juego nada que tenga que ver con el tiempo ni con la visibilidad. El ajedrez no es simplemente una simulación de la guerra, sino que es una abstracción de ella que captura un conjunto determinado de elementos. Estas características esenciales son las que proporcionan las similitudes de forma. Las diferentes transformaciones de lo viviente también pueden estar relacionadas a través de la historia o de la forma. Se cree que la vida en la Tierra surgió hace unos 3.800 millones de años, y que hace unos 3.500 millones de años nuestro planeta estaba poblado por una colección heterogénea de organismos unicelulares[4]. En este punto de la evolución no había organismos pluricelulares complejos, que evolucionaron más tarde, durante los últimos mil millones de años más o menos, mediante el desarrollo a partir de huevos fecundados. El desarrollo (la transformación de los cigotos en organismos pluricelulares) apareció mucho después de que comenzara la evolución, del mismo modo que el ajedrez surge muchos años después de que se comenzasen a librar batallas.
De igual forma, el aprendizaje surgió después del desarrollo. Los primeros organismos pluricelulares que se desarrollaron en nuestro planeta apenas eran capaces de aprender: no tenían cerebros que pudieran capturar relaciones nuevas en su entorno. Los sistemas nerviosos complejos comenzaron a evolucionar más tarde gracias a la aparición de cambios en la manera de desarrollarse de los embriones. Parte del desarrollo de algunos organismos se dedicó a la formación de cerebros con vías nerviosas de conexión, con lo que los seres vivos adquirieron la capacidad de aprender de su entorno, una capacidad que en la actualidad comparten muchos de nuestros primos, desde las babosas y los perros hasta los chimpancés. El aprendizaje vino después del desarrollo, al igual que el desarrollo vino después de la evolución.
El último tipo de transformación de lo viviente que surgió fue el cambio cultural. A medida que los humanos se diseminaban por la Tierra en grupos sociales, aprendiendo a domesticar y explotar a otras especies para su propio beneficio, generaban un excedente de comida que, junto con la capacidad humana para aprender e innovar, permitió que las sociedades mantuvieran y desarrollaran un abanico de especializaciones humanas, como por ejemplo constructores, soldados, artistas, instructores y administradores, lo que condujo a la formación de sistemas culturales elaborados. Las civilizaciones comenzaron a surgir tan solo en los últimos diez mil años, mucho después de que apareciera la capacidad para aprender. Por lo tanto, el cambio cultural es mucho más reciente que los otros procesos.
Las relaciones históricas parecen claras: primero vino la evolución, luego el desarrollo, después el aprendizaje y al final el cambio cultural. Esta secuencia temporal es la consecuencia de que cada proceso dependa de su predecesor. La capacidad de que los cigotos acaben produciendo organismos pluricelulares apareció gracias a la evolución. La capacidad de aprender depende del desarrollo previo de un sistema nervioso complejo dentro de un embrión. Y el cambio cultural solo resulta posible gracias a la capacidad de aprendizaje de los humanos. La cadena histórica no es más que el reflejo de una secuencia de dependencia.
Mientras que las relaciones históricas no parecen complicadas, las cosas son mucho más espinosas cuando acudimos a las relaciones de forma. ¿Existen similitudes fundamentales entre las distintas transformaciones, o cada proceso funciona de acuerdo con principios totalmente diferentes? En el pasado se han intentado identificar con regularidad las similitudes de forma sin ningún éxito, en gran parte debido a dos tipos principales de confusiones.
Uno de los errores tiene que ver con confundir lo familiar con lo fundamental. Como humanos, estamos familiarizados con la idea de diseñar y fabricar cosas, como ropa, muebles y casas. Por lo tanto, no parece natural utilizar la idea de fabricar como modelo general de cómo ocurren las transformaciones de lo viviente. Un árbol o una rana se podrían construir del mismo modo que un relojero fabrica un reloj, pero en vez de un humano, necesitaríamos que un hacedor divino más poderoso realizara el trabajo, aunque el principio de que un agente externo sea responsable del diseño y de la construcción sería el mismo. Esta idea de un agente externo o hacedor divino tiene una larga tradición y forma parte intrínseca de muchas religiones. Gracias a la capacidad retrospectiva de la ciencia podemos ver dónde radica el problema de esta explicación: la capacidad para diseñar y crear cosas que tenemos los humanos es un rasgo complejo que apareció mucho más tarde que la evolución. Utilizar la idea de fabricar como principio explicativo global no es más que intentar que un resultado complejo se explique a sí mismo. Caemos en esta trampa porque, como humanos, estamos muy familiarizados con la fabricación y no nos damos cuenta de que su complejidad descansa en toda una serie de transformaciones.
A los científicos les costó muchas generaciones derrotar esta equivocación. En ello resultó clave que Darwin identificara un mecanismo sencillo (la selección natural) capaz de explicar la diversidad de seres vivos de nuestro planeta. En vez de requerir un hacedor divino, la evolución de la vida avanza inexorablemente en respuesta al modo en el que los organismos se reproducen e interaccionan con su entorno. Su empeño por establecer este punto de vista dejó un legado duradero. Estableció una importante separación entre nuestros conceptos sobre las actividades humanas (diseño y creatividad) y el modo en el que reflexionamos sobre los procesos biológicos como la evolución. Son cosas muy diferentes que comparamos por nuestra cuenta y riesgo.
Regresemos al ajedrez para ilustrar otro tipo de error en la búsqueda de similitudes de forma. Fijémonos en la siguiente conversación entre un maestro de ajedrez y su pupilo principiante:
MAESTRO: Esta pieza se llama «caballo» porque puede saltar por encima de otras piezas.La equivocación tiene su origen en que el pupilo está tomando demasiado al pie de la letra la similitud entre la pieza de ajedrez y un caballo. Los caballos (con o sin jinete) tienen una relación meramente abstracta o simbólica. De hecho, se pueden aprender las reglas del ajedrez sin mencionar nunca los caballos, simplemente basta con aprender la manera en que se mueve. El juego no se vería afectado en absoluto si los caballos fueran teteras, ya que lo que realmente importa es la naturaleza de los movimientos que realiza. Las comparaciones entre los caballos con sus jinetes, si se toma de forma demasiado literal, acabarán distrayendo, llevando a una analogía errónea que confundirá más de lo que ayuda.
PUPILO: Entonces lo hará saltar el jinete, ¿no?
MAESTRO: No lo hace saltar el jinete, sino el jugador.
PUPILO: ¿Cómo lo consigue?
MAESTRO: Cogiendo el caballo y elevándolo por encima de las otras piezas.
PUPILO: Pero si el jugador lo hace saltar, ¿para qué le hace falta el jinete?
Este tipo de confusión aparece continuamente al comparar diferentes tipos de transformaciones de lo viviente. Un ejemplo sería la idea de Ernest Haeckel de que un huevo fecundado recrea su historia evolutiva a medida que se desarrolla. No hay duda de que la forma en que un huevo se desarrolla está vinculada con la evolución, pues gracias a esta surgió el desarrollo biológico. Pero al tratar de llevar esta relación demasiado lejos, esto es, que el desarrollo repita literalmente las etapas de la evolución, Haeckel acabó errando el tiro: no alcanzó el nivel de abstracción correcto y se lió haciendo comparaciones falsas y estériles. Las ideas de Edelman de que el cerebro funciona de acuerdo con los principios de Darwin han sufrido críticas similares por intentar aproximar demasiado dos procesos que están muy distantes.
Para determinar los principios comunes necesitamos trabajar en el nivel de abstracción correcto, que solo se alcanzará si se tiene un conocimiento razonable de lo que se quiere comparar. La comparación entre la guerra y el ajedrez solo dará algo con sentido después de que comprendamos lo que entraña cada uno. No hace falta que seamos expertos ajedrecistas ni que sepamos comandar un ejército, sino que debemos tener una idea global del funcionamiento de los juegos de mesa y de las batallas. Así podremos conocer tanto sus similitudes como sus diferencias. La guerra y el ajedrez son territoriales, si bien hay que saber que existen numerosas diferencias en el significado de territorio para cada caso: en la guerra es el terreno, mientras que en el ajedrez se refiere a una región del tablero.
De igual modo, para extraer los principios comunes que subyacen en las transformaciones que afectan a lo viviente necesitamos un conocimiento amplio de cómo funcionan. Esto se conoce desde hace muy poco gracias a los avances del conocimiento científico, en particular en los campos del desarrollo y del aprendizaje. Las comparaciones anteriores se habían visto obstaculizadas por la falta de conocimientos sobre cada proceso, lo que conducía a confundir lo familiar con lo fundamental, o a realizar abstracciones equivocadas. ¿Cómo deberíamos abordar ahora este problema?
§. Receta creativa para la vida
El crítico de arte chino del siglo VI Xie He sacó a relucir los seis ingredientes que pensaba que eran importantes para definir la calidad de una pintura[5]. Traducidos grosso modo, se trata de vitalidad, pinceladas, forma natural, colores, composición y copia. Los seis aspectos (que no son completamente independientes entre sí), con la excepción del color, se ilustran en Caquis (figura 1), un cuadro del monje del siglo XIII Mu Qi. La vitalidad de esta pintura procede en parte de la pincelada viva. De igual forma, la composición depende de la disposición de las frutas que se copiaron. En lugar de proporcionar un conjunto de características independientes, Xie He resaltó la interacción de algunos ingredientes clave que pensaba que ayudarían a apreciar el cuadro. Otros seguramente propondrán una lista de ingredientes diferente, por lo que las elecciones de Xie He nos gustarán o no en función de lo que nos ayuden a organizar nuestro conocimiento.Lo mismo ocurre con la manera en que organizamos nuestras ideas. Existen muchas formas de presentar nuestro conocimiento de procesos como la evolución, el desarrollo, el aprendizaje y el cambio cultural. En este libro presentaré un punto de vista muy particular que pone el énfasis en algunas características predominantes. Resaltaré siete principios fundamentales que subyacen a los procesos desde la evolución de las bacterias hasta el funcionamiento de nuestro cerebro. No piense que hay algo misterioso con el número siete, simplemente que los principios se han agrupado de forma natural en siete categorías, igual que Xie He encontró seis categorías para definir la pintura.

Figura 1. Caquis, Mu Qi (activo en 1269). Daitokuji, Kioto.
Me ha costado varios años llegar a este punto de vista. Comencé a pensar en estos temas cuando escribía un libro anterior,El arte de los génesis[6], que se centraba en el desarrollo, pero que también daba unas pinceladas sobre otros procesos, como la evolución y el arte. Relacioné estos procesos con el desarrollo a través de la historia o de la analogía. Pero había tantas similitudes que me hizo pensar en la posibilidad de que existieran unas conexiones más fundamentales. Estas ideas comenzaron a cristalizar en cuanto acabé de escribir el libro y me familiaricé con el pensamiento informático guiado por mis investigaciones científicas. Cayó en mis manos un ensayo revelador escrito en 1990 por el informático Christoph von der Malsburg, en el que identificaba tres principios básicos que eran comunes para todos los sistemas autoorganizados[7]. Hacía mucho hincapié en el funcionamiento del cerebro y su desarrollo, pero también se daba cuenta de la importancia que tenían estos principios para la evolución. La lectura de sus ideas me hizo mirar de otra forma los fundamentos del pensamiento biológico, en particular a la luz de lo que se había descubierto en los últimos años.
Me puse a reducir al mínimo las teorías de cada campo de la biología hasta dejar al descubierto los principios esenciales y entonces volví a examinarlos con aires renovados, buscando lo que podrían tener en común. Por supuesto, cuando uno busca con suficiente ahínco siempre acaba encontrando trivialidades comunes entre cualquier conjunto de procesos. Lo sorprendente del resultado fue, sin embargo, que los puntos de encuentro que aparecieron no eran superficiales, sino que iban a la esencia de cada proceso. Me sirvieron para definir las interacciones principales que en cada caso llevan a las transformaciones. Conseguí ver las transformaciones vivientes de una manera más profunda y más unificada, como diferentes manifestaciones de una receta creativa común.
Para dar a conocer el significado de la receta creativa para la vida necesitamos conocer cómo se aplica a cada tipo de transformación de lo vivo. Comenzaremos con la evolución (capítulos 1 y 2), que es la madre de todas las otras transformaciones y nos proporciona nuestra primera visión libre de obstáculos de cómo funciona la receta creativa para la vida. Entonces echaremos un vistazo al desarrollo, a cómo un cigoto microscópico se acaba convirtiendo en un árbol o un niño (capítulos 3 a 5). Hasta los años ochenta del siglo XX no empezamos a conocer bien cómo ocurría. Al recurrir a este conocimiento, veremos que a pesar de las muchas diferencias existentes, los principios del desarrollo muestran algunas similitudes sorprendentes con la evolución: se trata de la misma receta creativa, pero vestida con otro traje. Entonces pasaremos al aprendizaje, primero contemplando cómo organismos como las babosas y las hierbas responden a los cambios de su entorno (capítulo 6). Estos mecanismos de respuesta básicos nos proporcionarán algunos ingredientes fundamentales para el aprendizaje. El examen del funcionamiento del aprendizaje nos mostrará que se basa en la misma receta creativa que la evolución y que el desarrollo (capítulo 7), la misma receta que nos ayuda a entender la forma en que los animales aprenden a actuar, reconocerse e interpretar su entorno, proporcionándoles la base de la inteligencia y de la creatividad humanas (capítulos 8 y 9). Todos los casos de aprendizaje están impregnados de evolución y desarrollo, y nos muestran la manera en que una forma de la receta creativa para la vida puede enmarcar a otra (capítulo 10).
Entonces nos dirigiremos al cambio cultural (capítulos 11 y 12). Esta es posiblemente la transformación viviente más compleja, aunque paradójicamente es la que nos parece más familiar al ser nosotros mismos participantes activos de la sociedad. Podríamos identificar muchos factores perfectamente definidos que intervienen en el cambio cultural, como la creatividad humana, las personas carismáticas, las luchas de poder, el desarrollo económico y los cambios del medio ambiente. Es el ámbito de la historia, de la sociología y de la economía, y podría parecernos que hay poco que aprender de la biología. Pero la receta creativa para la vida permite contemplar el cambio cultural desde una perspectiva más amplia: en lugar de verlo como un proceso aislado que separa a los humanos del resto de los animales mediante un abismo infranqueable, veremos que está conectado con otros procesos tanto en forma como en historia. El cambio cultural es la cuarta manifestación de la receta creativa para la vida, que se fundamenta en las otras tres y las incorpora.
Transmitir este punto de vista requiere que se tengan unas amplias miras al tiempo que se ofrecen explicaciones científicas rigurosas. Esto significa que no he eludido dar detalles científicos cuando era necesario, aunque siempre he procurado que las explicaciones resulten comprensibles para la mayor parte del público. Animo a los lectores con pocos o ningún conocimiento científico a que sigan leyendo si encuentran pasajes complicados. Espero que quienes tengan más formación científica aprendan mucho de este libro al contemplar los campos de la ciencia tratados desde un punto de vista nuevo y unificador. Algunos pensarán que los campos que le son familiares están tratados de una forma muy selectiva, algo inevitable cuando se contemplan muchas disciplinas y las conexiones entre ellas, por lo que espero que estos lectores me perdonen las muchas omisiones.
Para ayudar a transmitir mi punto de vista, no mantengo la creatividad humana lejos de la narración científica, sino que recurro a ella constantemente a lo largo del libro. En muchas ocasiones utilizo pinturas para ilustrar los principios, temas o ideas que toco porque nos ofrecen puntos de acceso visuales y también sirven para recordarnos las muchas perspectivas desde las que podemos observar las cosas. Los temas científicos y artísticos acaban apareciendo juntos al final del libro. Veremos entonces que la evolución, el desarrollo, el aprendizaje y la cultura forman un gran ciclo, una serie de transformaciones relacionadas mediante las cuales la receta creativa para la vida acaba volviendo la vista sobre sí misma.Contenido:
§. Principio de la variabilidad en la poblaciónLas manzanas que comemos hoy en día no son exactamente iguales a las silvestres. Se cree que todos los manzanos cultivados descienden de poblaciones naturales de Malus pumila de la región Tian Shan en el Asia Central[8], cuya fruta se comen los grandes mamíferos, como los osos, y dispersan las semillas por las heces. A lo largo de muchas generaciones de cultivo y selección por los humanos, los manzanos silvestres se han transformado para producir una fruta más adaptada a nuestro gusto y a nuestra mesa. Darwin propuso que el origen de todas las formas de vida residiría en un proceso análogo de descendencia con modificaciones. Pero mientras que el cultivo de manzanos depende de la selección artificial ejercida por los humanos, Darwin identificó una forma de selección sin intervención humana: la selección natural.
§. Principio de persistencia
§. Principio de refuerzo
§. Principio de competencia
§. Combinación de los principios
Los manzanos producen muchísimas más manzanas y semillas de las que conseguirán convertirse en árboles frutales maduros. Por lo tanto, las semillas compiten continuamente entre sí en la naturaleza por llegar al estado adulto. Supongamos que, entre una población silvestre de manzanos, algunos árboles, digamos que el 1% de la población, producen manzanas más atractivas para los osos. Quizá contengan algo más de azúcar, por lo que sabrían más dulces. Como a los osos les gustan más estas manzanas, serán las preferidas de su dieta y las que dispersarían por el bosque. Por lo tanto, cabría esperar que las semillas de las manzanas apetitosas acaben dando más manzanos en la generación siguiente. Si el rasgo apetitoso se hereda, la proporción global de árboles que producen estas manzanas dulces se incrementará, digamos, un 2%. Al cabo de varias generaciones, la proporción continuará subiendo hasta que la población esté repleta de árboles que produzcan las manzanas apetitosas que los osos diseminarían con más eficacia. Gracias a la selección natural, los manzanos estarán mejor adaptados que antes a la reproducción en su entorno.
Los escenarios de este tipo proporcionan una visión general del funcionamiento de la selección natural, pero también plantean muchos interrogantes. El proceso a menudo se limita a la supervivencia del más apto. Pero ¿qué queremos decir con el más apto? ¿Nos referimos a los que finalmente sobrevivirán y desplazarán a los demás, como los árboles con las manzanas más apetitosas? Si es así, ¿se reduce la selección natural a un razonamiento circular (la supervivencia de los que sobreviven)? De igual forma, ¿por qué los organismos suelen producir muchas más semillas o descendencia de la que luego conseguirá sobrevivir? ¿Por qué no producen la cantidad que sobrevivirá hasta la edad adulta? Finalmente, ¿de dónde procede la variabilidad de los rasgos, como el buen sabor de las manzanas? Para responder estas preguntas de un modo satisfactorio debemos conocer más a fondo el funcionamiento de la evolución.
La descripción tradicional de la evolución mediante la selección natural recurre a tres principios esenciales[9]. El primero es que cada individuo de una especie es diferente de los demás. El segundo es que parte de las diferencias individuales se consiguen heredar y pasan de una generación a otra. El tercero es que los organismos se multiplican a más velocidad que la capacidad del entorno, con el resultado inevitable de que muchos morirán. La selección natural no es más que la consecuencia de la acción simultánea de estos tres elementos.
La explicación de la selección natural y de la evolución que ofrezco aquí se desvía un poco de esta descripción basada en tan solo tres elementos. En su lugar, describo el proceso mediante siete principios, y a veces utilizo términos que no significan exactamente lo mismo que en su uso tradicional. La razón por la que adopto este planteamiento tan peculiar es que mi objetivo global no es solo describir el funcionamiento de la evolución, sino también resaltar los ingredientes fundamentales que comparte con el desarrollo, el aprendizaje y el cambio cultural. Cuando queremos comprender la esencia de un proceso, suele resultar útil identificar los puntos de encuentro. Consideremos las transiciones del agua mencionadas en el capítulo anterior. Si estudiamos cómo hierve, podríamos deducir que lo que está pasando es que, a medida que se añade energía al sistema por calentamiento, las moléculas de agua adquieren más energía y movimiento hasta que alcanzan un punto en el que se liberan y se expanden para formar el vapor. La misma explicación se aplicaría a la fusión del hielo, salvo que el agua no se expande durante esta transición, sino que se contrae gracias a que sus moléculas en el hielo se mantienen en una configuración abierta, que se colapsa sobre sí misma cuando se liberan durante la fusión. Por lo tanto, el hielo flota en el agua al ser menos denso que el líquido que tiene debajo. Resulta más bien insólito que un líquido sea más denso que la forma sólida de una sustancia, puesto que la mayoría de los sólidos se vuelven menos densos cuando se funden. No obstante, este comportamiento excepcional del agua resulta informativo porque nos dice que la expansión durante el calentamiento no es fundamental para los cambios de estado de la materia[10]. Vemos que lo que realmente cuenta es que las moléculas ganan energía y adquieren más libertad de movimiento, y estas son las características comunes a las diferentes transiciones. La identificación de los elementos comunes nos ayudará a llegar hasta el fondo de lo que está sucediendo, a diferencia de lo que ocurre con lo que resulta más circunstancial.
Por supuesto, existe el peligro de que este planteamiento pueda conducirnos a identificar elementos comunes falsos o superficiales. Al comparar la fusión del hielo con la evaporación del agua podríamos advertir un cambio de aspecto en ambas transiciones: el agua líquida no se parece en nada al hielo y el vapor es muy distinto del agua líquida. Pero mientras que ambas transiciones comparten un cambio de aspecto, sería difícil considerar que esta característica común constituya un principio explicativo, sino más bien una descripción porque el cambio de aspecto no nos da ninguna clase de pista sobre el mecanismo de lo que ocurre. De igual forma, al determinar los principios que aunarían las transformaciones de lo viviente, es importante no buscar elementos comunes solo porque nos interese encontrarlos. El objetivo consiste en buscar los elementos mecánicos y las interacciones que sean similares, si existieran. Desde luego, no estaría escribiendo esto sin el convencimiento de que tales elementos existen y de que pueden ayudarnos a conocer más a fondo lo que conllevan las transformaciones en los seres vivos.
Para presentar los elementos comunes con más claridad utilizo una serie de términos aplicables a todas las transformaciones de lo viviente. Esto quiere decir que algunos temas se colocarán bajo epígrafes ligeramente inusuales. Por ejemplo, en este capítulo, el tema de la transmisión hereditaria entra dentro del principio de persistencia, y el tema de la multiplicación de los organismos entra dentro del principio de refuerzo. El nombre de cada uno de estos principios tiene un significado general y otro específico, al igual que un concepto como «composición» tiene un significado amplio y otro particular. Trataremos de la composición con respecto a muchas cosas, como cuadros, música o poesía. En todas ellas se hará referencia a una organización global, pero lo que se ordene será específico en cada caso (los colores en un lienzo, las notas musicales o una serie de palabras). De igual forma, veremos que cada uno de los principios que encontramos para las transformaciones de lo viviente tiene un significado general y uno específico.
Al buscar los fundamentos comunes también distingo algunos componentes que muchas veces se estudian juntos, con lo que aclaro la función de cada uno y expongo los principios que algunas veces se ignoran o se dan por sabidos. En este capítulo, por ejemplo, la idea de que los organismos se reproducen a un ritmo superior a la capacidad del entorno se subdivide en dos principios que se influyen mutuamente: refuerzo y competencia. En el siguiente capítulo nos toparemos con principios como la riqueza combinatoria y la recurrencia, que no se suelen incluir de forma explícita entre los ingredientes de la evolución. El lector no encontrará en ningún otro sitio esta colección de siete ingredientes que interaccionan, lo que denomino la receta creativa para la vida, para explicar las transformaciones de los seres vivos. Esta novedad no implica que vaya a presentar ninguna nueva teoría explicativa para determinados casos, sino que los principios e interacciones que describo han surgido al verlos desde una perspectiva que tiene en cuenta todas las transformaciones a la vez, y no por separado.
Para apreciar lo general, primero debemos conocer lo específico. Dado que la evolución es la madre de todas las otras transformaciones de lo viviente, resulta apropiado comenzar con ella a modo de ejemplo inicial de la receta creativa para la vida. En este capítulo explico los primeros cuatro principios de la receta porque proporcionan el núcleo de la selección natural. Los otros tres principios se tratarán en el capítulo siguiente.
§. Principio de la variabilidad en la población
Nunca estaremos seguros de si ganaremos a la ruleta. Si apostamos al rojo, cabría esperar que ganemos aproximadamente la mitad de las veces, pero no podemos predecir el resultado de ninguna apuesta en concreto. La probabilidad de ganar es en realidad ligeramente inferior a la mitad porque, además de las 18 casillas rojas y de las 18 negras que tiene el plato de la ruleta, hay una casilla verde, y si la bola cae en ella, la banca gana la apuesta (las mesas de ruleta estadounidenses tienen dos casillas verdes, por lo que la probabilidad de ganar es ligeramente inferior). Al cabo de muchas jugadas, tenderemos a perder al apostar al rojo porque la probabilidad de ganar es de tan solo 18/37, que está justo por debajo del 50%. Estos pequeños sesgos hacen que los casinos sean rentables, porque se basan en la estadística.El razonamiento estadístico conlleva dos elementos. Uno es el concepto de variabilidad: existe un abanico de posibilidades para cada jugada de la ruleta y la variabilidad proporciona la fuerza impulsora del juego. Esta clase de variabilidad se denomina a veces aleatoria debido a que resulta imposible predecir cada suceso por separado. Pero esto no significa que pueda pasar cualquier cosa: las bolas no caerán en ninguna casilla azul ni naranja del plato de la ruleta porque no existen tales colores. De igual forma, cuando echamos una moneda al aire hablamos de cara o cruz, no de negro o rojo. La variabilidad siempre se produce en un contexto determinado.
Además de la variabilidad, el otro elemento del razonamiento estadístico es el concepto de población. Aunque no seamos capaces de predecir lo que ocurrirá con cada apuesta a la ruleta, sabemos cuál será la tasa de ganancia o pérdida al cabo de muchas jugadas. Al considerar muchos sucesos juntos (una población), las peculiaridades empiezan a aflorar, como la tasa global de ganancias o pérdidas. La idea de una población supone que muchos sucesos están conectados de algún modo. Los resultados cuando se hace girar el plato de la ruleta con 38 casillas no pertenecerán a la misma población que los de un plato de 37. Al calcular probabilidades, tampoco mezclaríamos los números obtenidos de lanzar dados con los números obtenidos al girar un plato de ruleta. Cada población está basada en cierta clase de criterios unificadores.
El razonamiento estadístico comporta la relación de dos tipos de unidades: el individuo y la población. Se puede pensar que representan diferentes escalas o niveles: cuando nos acercamos, vemos la variabilidad de los sucesos (cada jugada a la ruleta); cuando nos alejamos, apreciamos el comportamiento de la población como un todo (la tasa total de pérdidas o ganancias). Gracias a la estadística relacionaremos los sucesos de un nivel con los del otro. Podemos observar el comportamiento de la población como el resultado colectivo de variaciones a nivel individual. De hecho, la estadística proporciona una de las herramientas más poderosas que tenemos para conectar los diferentes niveles o escalas, para acercarnos y alejarnos de un problema.
La selección natural también es un proceso estadístico en el que intervienen los conceptos de variabilidad y población. En este caso, la variabilidad tiene que ver con diferencias entre individuos. Siguiendo con el marco hipotético descrito antes, unos árboles proporcionan manzanas más dulces y otros menos deliciosas. La población es un grupo de individuos, la colección de manzanos que pueden reproducirse y cruzarse en un bosque. La selección natural nos dice cómo surgirían los sucesos a nivel de población (un incremento de la frecuencia de árboles con manzanas apetitosas) gracias al efecto acumulativo de las variaciones a nivel individual (el éxito reproductor de los diferentes manzanos). Estudiemos primero el origen de esta variabilidad.
Las variaciones hereditarias se encuentran en el ADN, una molécula larga formada por dos hebras en espiral. Hay cuatro tipos de unidades, denominadas bases nitrogenadas, a lo largo de cada hebra del ADN, o cuatro letras en el alfabeto del ADN, que se escriben habitualmente como A, T, G y C (por adenina, timina, guanina y citosina). El ADN de una planta o animal se denomina genoma y comprende miles de millones de tales bases concatenadas una detrás de otra para dar una secuencia. Igual que un gran trozo de texto se subdivide en palabras, cada genoma se subdivide en tramos de ADN denominados genes. Los humanos, por ejemplo, tenemos unos 25.000 genes en el genoma. Cada gen contribuye a determinadas características de un individuo, como el sabor de las manzanas o el color de los ojos.
A veces se producen ligeros cambios en la secuencia del ADN, como que una C sea reemplazada por una T en una posición determinada. Las mutaciones de esta clase tendrían su origen en la radiación absorbida por el ADN o en los errores que se producen durante la replicación de esta molécula. Si en el ADN de una célula que da lugar a los óvulos o a los espermatozoides aparece una mutación, pasará a la generación siguiente, con lo que se introducirá un gen mutante en la población. Como las particularidades de los individuos dependen de la secuencia de su ADN, esto ocasionaría variaciones hereditarias en la población, como la variabilidad del dulzor de una manzana o del color de los ojos.
No obstante, existen límites al tipo de variabilidad esperable. No existirán mutaciones que den lugar a manzanas con ojos azules porque la variabilidad siempre surge en un contexto particular, en relación con una secuencia de ADN o genoma determinados. Para el genoma de un manzano, las mutaciones podrían hacer que produzca diferentes tipos de manzanas, pero nunca darán lugar a rasgos humanos como los ojos. La variabilidad está definida por el genoma y el organismo de partida, por el contexto.
No se puede predecir si una mutación aparecerá en una posición específica del ADN en un momento determinado. Sin embargo, en una población de muchas moléculas de ADN sí hablaremos de la probabilidad de que aparezca una mutación en una posición determinada. Esta probabilidad suele ser muy baja: alrededor de una por cada mil millones de divisiones celulares. Incluso así, como el ADN es tan largo y la población está formada por muchos individuos, siempre surgirán mutaciones, aunque cada una constituya un suceso infrecuente: surgen continuamente en las poblaciones y proporcionan la variabilidad genética.
La variabilidad genética se potencia no solo por las mutaciones, sino también por la reproducción sexual. El genoma no suele estar en una única pieza larga de ADN, sino que está dividido en fragmentos de diferente longitud denominados cromosomas. Para los organismos con reproducción sexual, estos cromosomas forman parejas, cada uno procedente de un progenitor durante la reproducción. Cualquiera de nosotros, por ejemplo, tiene 23 pares de cromosomas en el genoma: de nuestro padre heredamos 23 y los otros 23 de nuestra madre. Los pares se pueden barajar, como los naipes, de una generación a otra cuando se producen los espermatozoides o los óvulos, de resultas de lo cual los cromosomas se reúnen en nuevas combinaciones cuando se fertiliza el huevo, y se incrementa la variabilidad genética de la población. También pueden surgir nuevas combinaciones genéticas gracias a los intercambios entre los cromosomas porque pueden romperse y volverse a unir durante la producción de los espermatozoides o de los óvulos, de tal forma que una porción de un cromosoma se intercambia con la correspondiente porción de su homólogo. Este intercambio (denominado recombinación) incrementa aún más la variabilidad de la población.
Por lo tanto, se está generando variabilidad hereditaria continuamente, lo que constituye una peculiaridad básica de todos los organismos y proporciona el primer elemento del razonamiento estadístico. Ahora vamos al segundo elemento: el comportamiento de la población.
La selección natural depende de la variabilidad de una población formada por muchos individuos. Consideremos el ejemplo de los manzanos, en los que la proporción que produce manzanas dulces se incrementaba poco a poco porque los osos dispersaban sus semillas con más eficacia. Es importante aquí considerar qué le ocurre a la población en conjunto en vez de a cada individuo en particular. Por ejemplo, un determinado árbol con manzanas amargas puede dejar más descendencia que otro que las tiene dulces. Quizá el árbol de las amargas crece en una parcela particularmente fértil del bosque, o quizás se encuentra cerca de una osera. Sea cual sea la explicación, el éxito de este árbol en concreto no es capital para la selección natural porque lo que cuenta es su comportamiento reproductor con respecto a la población completa. En el conjunto de la población, los árboles con manzanas dulces tienen de media más éxito reproductor que los de las amargas, lo que conduce a un incremento de la proporción de árboles con manzanas sabrosas. De igual forma, una determinada tasa de ganancias o pérdidas a la ruleta surge de un grupo (población) de apuestas, no de ninguna apuesta en particular. La selección natural actúa sobre la tasa de supervivencia y de reproducción de una población, y no sobre el destino de uno o dos individuos. Se trata de un proceso estadístico basado en la variabilidad de la población.
§. Principio de persistencia
El jueves 12 de septiembre de 1940, cuatro chicos franceses del pueblo de Montignac estaban explorando el agujero que había dejado al descubierto un árbol caído en los alrededores. Habían traído una linterna porque corría el rumor de que el agujero conducía a un pasaje subterráneo. Después de precipitarse al interior y alumbrar con la linterna, descubrieron una serie de cuevas subterráneas con las paredes cubiertas de pinturas de animales. Habían tropezado con una de las decoraciones más bellas del arte rupestre de la cueva de Lascaux. Las pinturas habían sobrevivido en un estado increíblemente bueno durante más de diez mil años (figura 2).La supervivencia durante tanto tiempo de estas pinturas es un reflejo de la estabilidad de la roca y del pigmento. Se trata de un testimonio de la resistencia de las fuerzas moleculares que mantuvieron la roca cohesionada y la pintura adherida a su superficie. Sin embargo, incluso aunque se produjese una desgracia y la cueva de Lascaux quedase destruida, seguiríamos teniendo una idea del aspecto de las pinturas porque existe otra forma de persistencia. Los negativos y positivos fotográficos y las imágenes electrónicas de Lascaux proporcionan otra manera de conservar cuya persistencia también depende de las fuerzas moleculares: las que impiden que se disgreguen las moléculas en las fotografías o en los dispositivos electrónicos. Pero aquí aparece el beneficio añadido de la seguridad en los números, porque la información se puede copiar y propagar considerablemente. Por esta razón, millones de personas que nunca han visitado la cueva pueden, a pesar de todo, apreciar las pinturas gracias a las muchas fotografías e imágenes que existen.

Figura 2. Pintura rupestre de la cueva de Lascaux, Francia, 15.000-10.000 a. C.
La secuencia de ADN también persistirá gracias a la replicación (copia). El ADN de cada cromosoma está formado por dos hebras que se emparejan entre sí, del mismo modo que los negativos y positivos fotográficos son complementarios. Siempre que hay una A en una hebra de ADN tiene que haber con una T en la hebra opuesta y formarán lo que se denomina un par de bases A-T. De igual forma, G y C están siempre una frente a la otra y forman el par de bases G-C. De este modo, la secuencia de bases de una hebra contiene toda la información necesaria para especificar la secuencia de la complementaria: la secuencia AGCT en una hebra determina que la otra sea TCGA, al igual que un negativo fotográfico siempre casa con su positivo. La replicación se lleva a cabo cuando las dos hebras del ADN se separan y cada una actúa como plantilla para especificar su complemento, lo que permite fabricar dos moléculas de ADN a partir de una: la hebra positiva especifica un nuevo homólogo negativo, mientras que la hebra negativa especifica un nuevo homólogo positivo. Esta copia tiene lugar cada vez que se produce la división celular para que cada célula hija termine con una copia del genoma. Con la copia del ADN también se consigue que la información se propague y pase de una generación a la siguiente.
Tal y como ya hemos aprendido del principio de variabilidad de la población, la secuencia de ADN no es completamente inalterable, pues se modifica por lesiones o mediante errores de copia. Si una mutación diera lugar a un organismo menos capaz de sobrevivir y de reproducirse, lo más probable es que la versión mutante se elimine de la población mediante la selección natural. Por otra parte, una mutación puede conducir en ocasiones a un mayor éxito reproductor, en cuyo caso la selección natural favorecería la diseminación de la versión mutante por la población. Y todo esto ocurre porque las variaciones son capaces de persistir gracias a la replicación.
Para que haya evolución tiene que existir un equilibrio entre la variabilidad y la persistencia. La variabilidad proporciona la materia prima para la selección natural, mientras que la persistencia permite que la información de una generación pase a la siguiente. La variabilidad sin la persistencia significaría que los cambios no se mantendrían ni se consolidarían durante la evolución. La persistencia sin la variabilidad llevaría al estancamiento de la evolución.
El equilibrio entre la persistencia y la variabilidad no es solo un requisito para la evolución, sino que es algo que ha evolucionado por sí mismo. La replicación del ADN, por ejemplo, depende de las proteínas que separan y copian sus hebras. Si estas proteínas funcionaran con negligencia y cometieran montones de errores cuando copian el ADN, se introducirían muchas mutaciones, lo que haría predominar la variabilidad sobre la persistencia. Sin embargo, la presencia de demasiados errores conduciría a que el ADN se copiase de forma ineficaz y su secuencia comenzaría a degenerarse, igual que una mala fotocopiadora va haciendo perder información de una imagen. Este impacto negativo de las copias negligentes significa que las proteínas responsables tenderían a estar desfavorecidas por la selección natural, lo que desplazaría el equilibrio a favor de la persistencia y en contra de la variabilidad. Durante miles de millones de años de evolución se ha producido una interacción continua entre la persistencia, la variabilidad y la selección. El equilibrio entre la variabilidad y la persistencia que vemos hoy en día en las poblaciones que nos rodean es el resultado de esta larga historia evolutiva.
La interacción entre la persistencia, la variabilidad y la selección proporciona el primero de los bucles de realimentación que nos encontraremos en este capítulo. Nos enseña que los diferentes principios que describo no son completamente independientes entre sí. La variabilidad y la persistencia no son solo ingredientes de la evolución, sino que se modifican mutuamente gracias a la evolución.
§. Principio de refuerzo
La variabilidad de la población y la persistencia hereditaria proporcionan el telón de fondo para la evolución, aunque por sí solas no proporcionan una fuerza impulsora para el cambio evolutivo. Tal fuerza emerge cuando consideramos una determinada clase de variabilidad: la que influye en la capacidad reproductora.Este proceso se basa en la ley del interés compuesto. Supongamos que tenemos una población de cien bolas que tienen la capacidad de duplicarse de vez en cuando, por lo que a veces pueden aparecer dos nuevas bolas a partir de una sola. Imaginemos que, durante una hora, estas duplicaciones conducen a que el número de bolas se incremente de media un 10%, lo que significa que, transcurrida una hora, tendremos unas 110 bolas. Ahora ya tenemos algunas bolas más con capacidad de duplicación. Por consiguiente, el número de bolas se habrá incrementado un poco más al cabo de otra hora, unas once bolas de media (el 10% de 110), lo que dará lugar a un total de 121 bolas. El número continuará incrementándose de este modo (según las reglas del interés compuesto) y, después de diez horas, tendremos unas 260 bolas. Después de cien horas, se habrán convertido en más de un millón.
Ahora supongamos que las bolas son de dos colores diferentes: negras y blancas. Podemos suponer que el color de la bola se transmite con fidelidad cuando esta se duplica, de manera que una bola negra las dará negras, y una blanca las dará blancas. Si comenzamos con el mismo número de bolas negras y blancas, y ambas se replican a la misma velocidad media, entonces la proporción permanecerá aproximadamente igual a medida que aumenta el tamaño de la población. Sin embargo, la situación será diferente si cada color se reproduce a distinto ritmo. Imaginemos que las bolas negras se incrementan a una velocidad media del 11% por hora mientras que las blancas lo hacen al 10%. Al comenzar con una población de 100 bolas negras y otras 100 blancas esperaremos terminar con unas 111 negras y otras 110 blancas después de una hora, de manera que las bolas negras constituirán algo más del 50% de la población (aproximadamente el 50,2%). Después de diez horas habrá unas 280 bolas negras y 260 blancas, por lo que la proporción de negras se habrá incrementado al 52%, más o menos. Después de cien horas, la población habrá crecido hasta dar millones, y la proporción de bolas negras será de un 70%. La ley del interés compuesto garantiza que la ligera diferencia del ritmo de reproducción entre las bolas negras y las blancas conducirá a una diferencia numérica que se refuerza hora tras hora, por lo que las bolas negras serán cada vez más numerosas que las blancas.
El refuerzo ocupa un lugar central en la selección natural. Los genes, o si hablamos con más propiedad, las versiones de los genes, que favorecen la reproducción de un organismo tenderán a estar mejor representados en la siguiente generación. Al ser solo un poco más abundantes, estas versiones génicas comienzan con una ventaja que se amplificará en la próxima generación. Al repetir este proceso una y otra vez, un gen que favorezca la reproducción reforzará continuamente su cantidad y se incrementará en la población.
Sin embargo, el refuerzo por sí solo no consigue que una versión de un gen reemplace a otra. Incluso si dejamos que nuestra población de bolas negras y blancas continúe creciendo durante muchos días, de modo que más del 99% sean negras, todavía encontraríamos un número inmenso de bolas blancas porque no se pierde ninguna bola, sino que simplemente tenemos cada vez más. El refuerzo hará incrementar la proporción, pero nunca sustituirá una versión del gen por otra. Para ver cómo se produciría el reemplazo, necesito introducir otro principio: la competencia.
§. Principio de competencia
La idea de la selección natural se le ocurrió a Darwin poco después de que volviera de sus cinco años de viaje a bordo del HMS Beagle, el barco en el que trabajó como naturalista. De resultas de este viaje, Darwin quedó convencido de que las especies no eran inmutables, sino que podían modificarse con el tiempo. Pero no sabía el mecanismo con el que explicar la manera en que las especies cambian y se adaptan. Entonces, en octubre de 1838, a los dos años de su regreso, estaba leyendo un libro de Thomas Malthus sobre el crecimiento de la población en el que señalaba que si la población humana seguía aumentando de acuerdo con las leyes del interés compuesto, la población mundial acabaría agotando los alimentos, a lo que seguirían guerras y hambruna. Esta idea puso el dedo en la llaga:
Me puse a leer el Primer ensayo sobre población de Malthus para entretenerme, y al estar bien preparado para apreciar la lucha por la existencia que domina por doquier gracias a la observación prolongada de las costumbres de los animales y de las plantas, de pronto fui consciente de que, bajo estas circunstancias, las variaciones favorables tenderían a conservarse y se perderían las desfavorables. El resultado sería la formación de especies nuevas. Al fin tenía una teoría con la que trabajar [11].
Darwin se dio cuenta de que si toda la progenie de las plantas o de los animales sobrevivía, el tamaño de la población sobrepasaría pronto los límites de lo que podría mantener el entorno. El resultado sería una continua lucha o competencia por la existencia, en la que morirían muchos individuos. Los mejor adaptados tendrían más posibilidades de sobrevivir, lo que daría lugar a una forma natural de selección. Leer a Malthus fue importante para Darwin porque le permitió unir dos ideas: las poblaciones tienden a crecer de acuerdo con la ley del interés compuesto, y este crecimiento se ve finalmente limitado porque los recursos del entorno son finitos. Tampoco fue por casualidad que Alfred Russel Wallace estuviera pensando en Malthus cuando le sobrevino la idea de la selección natural unos 20 años después de Darwin (pero antes de que Darwin lo publicara). En ambos casos, la confluencia de las ideas sobre el incremento reproductor y las limitaciones ambientales proporcionó la semilla para formular la selección natural.
El concepto de competencia suele venir ligado al de refuerzo porque, como advirtieron Darwin y Wallace, la competencia a menudo surge por la presión de los grandes números. Pero para entender mejor la selección natural, en primer lugar conviene separar estos dos conceptos y considerar lo que la competencia representa por sí misma, sin tener en cuenta el refuerzo. Esto nos ayudará a entender por qué la teoría de la selección natural no es un argumento circular sobre la supervivencia de los que sobreviven. Para apreciar los efectos de la competencia sin el refuerzo, volvamos a nuestro ejemplo de las bolas blancas y negras.
Imaginemos que un bombo de lotería contiene el mismo número de bolas blancas y negras, y que están mezclándose continuamente (figura 3, izquierda). Digamos que hay 500 bolas de cada color, por lo que nuestra población comprende mil bolas en conjunto. Cada cierto tiempo se extrae una muestra de unas pocas bolas por la boca del bombo. Después de un tiempo, se cierra la boca y obtenemos un determinado número de bolas, pongamos 100, a lo que llamaremos muestra poblacional. Como la muestra es menor que la población entera, podemos decir que las bolas del bombo de lotería están compitiendo entre sí por conseguir un lugar en la muestra. Al utilizar la palabra «competencia» aquí, no quiero decir que las bolas están peleándose entre sí, sino que hay un gran número de candidatas para un escaso número de sitios.
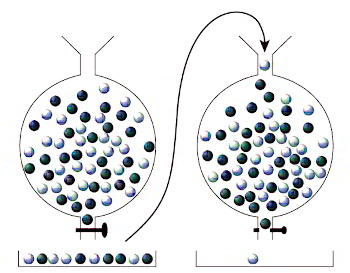
Figura 3. Muestreo de una población de bolas, reproducción de las bolas y nuevo muestreo.
Consideremos en primer lugar una situación en la que las bolas blancas y negras no muestran ninguna propiedad diferente, o sea, que compiten igual de bien por un sitio en la muestra (muy parecido a la competencia humana, en la que muchos candidatos con habilidades similares compiten por un número limitado de puestos). Si comenzamos con el mismo número de bolas blancas y negras, cabría esperar que la proporción se mantenga más o menos igual durante las generaciones. Pero sorprendentemente, después de unas 100 generaciones, la población contendrá, con mucha probabilidad, solo bolas blancas o solo bolas negras. Este resultado tiene que ver con la manera en que se acumulan las variaciones aleatorias. En la primera muestra de bolas, por ejemplo, podríamos no encontrar exactamente 50 bolas blancas y otras 50 negras, sino que podríamos encontrar más de uno de los colores. Supongamos que encontramos 53 bolas negras y 47 blancas. Después de la replicación, tendremos unas 530 bolas negras y 470 blancas para el siguiente bombo de lotería. La fluctuación de nuestra muestra se ha incorporado en la población de la lotería, lo que significa que la probabilidad de sacar una bola negra en la siguiente muestra es un poco mayor de un medio (53%). Las fluctuaciones de la muestra siguiente podrían realzar más o quizá disminuir esta diferencia. De esta forma, la proporción de bolas blancas y negras subirá o bajará, y las fluctuaciones se pasarán una y otra vez de un bombo al siguiente. Después de 100 generaciones de acumulación de fluctuaciones de este tipo, la población podría acabar derivando a un único color. Cuando esto ocurre, no hay vuelta atrás y la población se queda monocroma, en este caso, negra. Decimos entonces que la población ha quedado fijada en las bolas negras. Precisamente por el efecto de repetir el muestreo (se repite la competencia por un número limitado de sitios), la población ha evolucionado desde tener bolas blancas y negras a partes iguales, a acabar fijando un único color. Esto ocurre incluso aunque cualquier bola en particular, blanca o negra, tenga exactamente la misma probabilidad de ser muestreada en cada generación (cada participante en nuestra competición tiene exactamente la misma capacidad y, por tanto, la misma probabilidad de abrirse paso).
Comparemos este resultado con lo que ocurre si muestreamos una única vez. Si tuviéramos que muestrear 100 bolas de una mezcla con el mismo número de bolas blancas y negras, la probabilidad de que todas sean de un color es de aproximadamente 1 en 1030, o una en un quintillón. Si solo hubiera que coger muestras únicas, habría que esperar un tiempo superior a la edad del universo para tener la suerte de sacar todas las bolas del mismo color. En cambio, con nuestro esquema anterior de repetición de ciclos de muestreo y replicación, acabar con un color es casi inevitable y se tarda tan solo 100 generaciones (el número de generaciones necesarias es proporcional al tamaño de la población de la muestra). Al introducir una muestra limitada (competencia por un escaso número de sitios), y luego dejar que cada generación crezca a partir de la anterior, hemos cambiado el carácter del proceso, de manera que ahora hay una tendencia natural al reemplazo. Existe la misma probabilidad de que el color dominante sea blanco o negro, pero con seguridad, uno acabará reemplazando al otro.
La vida también es una lotería. Cuando los manzanos silvestres de los bosques de Tian Shan producen su enorme cantidad de manzanas, solo una pequeña parte de las semillas acabarán dando árboles maduros que producirán más semillas para la siguiente generación, simplemente porque la región de Tian Shan tiene un tamaño finito y solo es capaz de mantener unos pocos manzanos. Los árboles maduros equivalen a la muestra de población del bombo de lotería. Si imaginamos que cada árbol produce un millón de semillas por generación, entonces de media solo una entre un millón de ellas crecerá para dar un árbol. En este caso, la muestra poblacional sería la millonésima parte de la población del bombo, con lo que la competencia por los sitios es intensa. Al igual que el ciclo de la lotería que describimos anteriormente, el proceso se repite generación tras generación.
Las bolas blancas y negras son equivalentes a las diferentes versiones de un gen en la población de manzanos. Gracias a la mutación, continuamente se introduce en las poblaciones una variabilidad genética de este tipo. A medida que surgen nuevas versiones, algunas tienden a derivar hacia un número mayor por el repetido muestreo (competencia) que se produce con cada generación. Esta deriva genética que se debe a los límites en el tamaño de la población se produce incluso si la versión del gen no altera ni la supervivencia de la semilla ni su reproducción. Con el tiempo, algunas variaciones se quedan fijas en la población, lo mismo que vimos con el continuo muestreo de bolas.
La deriva genética ilustra que la introducción de un límite en el tamaño de una población puede tener un efecto drástico sobre lo que ocurre al cabo de varias generaciones. En el apartado anterior sobre el principio de refuerzo vimos que la ley del interés compuesto conlleva un incremento paulatino de la proporción de una versión de un gen en caso de que tenga una ventaja reproductiva con respecto a otra. Sin embargo, como la otra versión no se eliminaba, la nueva nunca llegaba a sustituirla completamente. Al introducir un límite en nuestra población reproductora, algunas variantes de genes pueden alcanzar la supremacía total, incluso si las versiones competidoras del gen tienen la misma tasa reproductora. La competencia por sí sola, sin tener en cuenta los méritos de cada versión, permite que una población evolucione y cambie con el tiempo. Ahora podemos ver por qué la teoría de la selección natural no es un argumento circular. Ya hemos visto que una forma puede sobrevivir y reemplazar a otra en una población simplemente por el efecto de repetir el muestreo (deriva genética). La importancia de esto se manifiesta en que una población es capaz de evolucionar, con el consiguiente cambio de su acervo genético, en ausencia de selección natural. La evolución y la selección natural no son lo mismo, pues cuando se trata la selección natural, el más apto no designa a los que acabarán sobreviviendo en esta población. Si así fuera, equivaldría a afirmar que la bola negra es más apta que la blanca a pesar de que su abundancia se deba a la repetición del muestreo, sin ninguna ventaja inherente sobre la blanca. La eficacia biológica o fitness tiene un significado completamente diferente, ya que no se refiere a los que finalmente sobrevivirán, sino a la introducción de un sesgo en la competencia, como veremos ahora.
§. Combinación de los principios
Hasta ahora hemos supuesto que las bolas blancas y negras de nuestro bombo de lotería tienen la misma capacidad de reproducirse y competir por un sitio en la población de muestra, por lo que tienen la misma probabilidad de acabar invadiendo la población. Veamos lo que pasa cuando introducimos alguna diferencia entre sus propiedades. Supongamos, por ejemplo, que las bolas negras se replican más rápido que las blancas. Se trata de la misma situación que vimos en el principio de refuerzo, pero ahora la combinamos con la idea de un bombo de lotería con pocos sitios en la muestra (competencia). Después de tomar la muestra, la mayor capacidad reproductora de las bolas negras hace que su cantidad se incremente más que la de las blancas. Por lo tanto, es más probable que contribuyan más a la siguiente generación y se cree un sesgo a favor de las negras, sesgo que se reforzará con cada generación. Al cabo de varias rondas de muestreo, es más probable que en la población se fijen las negras y no las blancas.Al combinar el refuerzo (mayor tasa reproductora de las bolas negras) con la competencia, están pasando dos cosas. Primero, ahora es más probable que en la población se fijen las bolas negras y no las blancas: el sistema se ha vuelto sesgado hacia el color que se reproduce mejor. Segundo, la proporción de bolas negras se incrementa más rápido que si los dos colores tuvieran la misma capacidad reproductora porque su proporción se ve estimulada con cada generación, lo que hace incrementar el ritmo de crecimiento del porcentaje de bolas negras. La velocidad a la que se incrementa la proporción de bolas negras acaba decayendo a medida que se hacen más predominantes en el bombo porque a medida que la proporción de bolas blancas disminuye, las negras comienzan a competir cada vez más entre sí por conseguir un sitio en la muestra. A la larga, el éxito de las bolas negras reduce su tasa de incremento. A pesar de todo, las bolas negras tardan menos en fijarse que cuando no tenían ninguna ventaja sobre las blancas. Estas dos consecuencias (resultado sesgado y menor tiempo de fijación) las suelen utilizar los biólogos evolucionistas para inferir que en una población se produce selección natural, y no solo una simple deriva genética.
Hay otra manera de favorecer la suerte de las bolas negras. Supongamos que pesan un poco más que las blancas y que tienden a estar más cerca del fondo del bombo, por lo que aumentará su probabilidad de caer en la muestra. Las bolas negras tendrán ventaja eficaz sobre las blancas a la hora de colocarse en la población de muestra (de manera parecida a la competencia humana, en la que los candidatos no son igualmente capaces, sino que algunos son mejores que otros y tienen una mayor probabilidad de ganar). Que haya más bolas negras en la muestra significa que contribuyen más a la siguiente generación, incluso aunque se repliquen al mismo ritmo que las blancas. La ventaja de las negras se refuerza con cada generación, por lo que es más probable que acaben colonizando el bombo entero. Un cambio en la capacidad para hacerse sitio en la muestra tiene un efecto similar que cambiar la tasa reproductiva. Se trata de otra forma de refuerzo, pero una que aparece durante el propio proceso de competencia.
A la población de manzanos le afectan los mismos principios: si una versión génica promueve la capacidad de supervivencia de las semillas para crecer y dar un árbol maduro, entonces dicha versión tenderá a incrementarse en la población, puesto que tiene una ventaja selectiva, una fitness más alta, al igual que la bola más pesada de nuestro bombo de lotería. Otro tipo de ventaja selectiva tiene lugar cuando la versión génica mejora la capacidad reproductiva del manzano: una versión que vuelva las flores de las manzanas más atractivas a las abejas, por ejemplo, podría hacer que se produjeran más semillas. Esta ventaja reproductiva también se reforzará en cada generación, aumentando su probabilidad de fijarse.
Estas dos formas de refuerzo, un incremento de la capacidad de sobrevivir y un incremento de la capacidad de reproducirse, marcan la diferencia entre la selección y la deriva genética. Con la deriva genética, las versiones de los genes tienen la misma probabilidad de quedarse fijadas, mientras que será distinta con la selección (se promueven las que favorecen la reproducción y el crecimiento). Esta es la manera en que la selección natural ajusta el organismo a su entorno: favorece las peculiaridades que incrementarán la reproducción y la supervivencia.
La interacción entre refuerzo y competencia forma el núcleo de la selección natural y se resume en el diagrama de la figura 4. El bucle de realimentación positiva representa el refuerzo: si una versión génica ayuda al éxito reproductor, tiende a reforzarse a sí misma y a aumentar su presencia en cada generación. El bucle de realimentación negativa representa la competencia y surge por las limitaciones ambientales que constriñen el tamaño de la población. Una consecuencia del límite poblacional es que la frecuencia de una versión génica aumenta a expensas de la otra, lo que finalmente conduce a la eliminación de la versión rival. Otra consecuencia del límite poblacional es que una versión génica compite contra sí misma a medida que se hace más predominante en la población y acaba siendo víctima de su propio éxito: a medida que prolifera en la población, es más probable que los individuos rivales tengan la misma versión del gen, por lo que disminuye su ritmo de aumento. Cuando la versión génica reemplaza por completo a su rival, todos los individuos tendrán la misma probabilidad de competir porque compartirán las mismas ventajas. El resultado global es que todos los individuos estarán más capacitados para sobrevivir y reproducirse. Una adaptación se habrá establecido en la población.
El doble bucle de realimentación entre refuerzo y competencia ayuda a explicar por qué los organismos suelen tener más descendencia de la que conseguirá sobrevivir. Si comenzamos con una población que produce el mismo número de descendientes que los que puede mantener el entorno, no habrá competencia porque todos podrán sobrevivir. Pero supongamos que por mutación surge una versión génica que incrementa el ritmo de reproducción. La versión génica es muy probable que se extienda por la población mediante el refuerzo (bucle positivo en la figura 4). A medida que esto ocurre, la población se reproduce con más eficacia y deja más descendencia que antes, por lo que la cifra total excederá los límites del entorno y acabará apareciendo la competencia por los recursos limitados (bucle negativo en la figura 4). El incremento del éxito reproductor conduce a la competencia que lo limita. Este doble bucle de realimentación no solo es capaz de impulsar al cambio evolutivo, sino que también lo mantiene. El doble bucle de realimentación por medio del refuerzo y la competencia es el motor en el núcleo de la selección natural. Se trata de un motor alimentado por un equilibrio entre la variabilidad y la persistencia poblacionales. Sin variabilidad no habría nada que mantuviera el motor en marcha, y sin persistencia no se conseguiría provocar un cambio. También hemos visto otro bucle de realimentación entre el motor y su combustible: la exactitud de la copia del ADN, y por ende el grado de variación, está modulada por la selección natural.
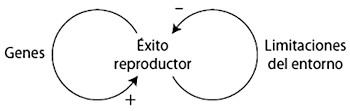
Figura 4. Interacción entre el refuerzo (bucle positivo) y la competencia (bucle negativo) en la selección natural.
Aunque estos bucles de realimentación están implícitos en muchas descripciones de la evolución, hemos podido verlos con más claridad después de apartar los ingredientes que los forman. Ahora podemos apreciar cómo la selección natural es impulsada mediante la interacción entre cuatro principios: variación en la población, persistencia, refuerzo y competencia. En los próximos capítulos veremos que en el centro de otros tipos de transformaciones de los seres vivos se encuentran unos principios y bucles de realimentación parecidos que nos proporcionan una forma común que surge con muy diferentes apariencias (igual que el ajedrez y la guerra tienen una forma común a pesar de que se diferencian en otros muchos aspectos). Pero, por ahora, quiero seguir describiendo la evolución atendiendo a algunas peculiaridades que no hemos tenido en cuenta o a las que no hemos dado importancia.
A lo largo de este capítulo hemos supuesto que la selección natural opera en un contexto determinado: una población de individuos. Se puede decir que la selección natural actúa dentro de la población de manzanos y que favorece a un tipo de manzanas en lugar de a otras; o que actúa dentro de la población de osos y favorece a unos osos en lugar de a otros. Pero nunca diremos que la selección natural favorece a un manzano más que a un oso porque, aunque haya más manzanos que osos, esto no significa que la selección natural favorezca más a unos que a otros. La selección natural se ocupa del éxito relativo de los individuos dentro de una misma población, no de su abundancia absoluta. Esto hace que nos preguntemos por qué existen individuos para empezar, y por qué se organizan en poblaciones tan variopintas. ¿Por qué unas poblaciones están formadas por muchos manzanos y otras solo por osos? Para responder a estas preguntas examinaremos los otros tres principios que faltan para completar la receta creativa para la vida.
Capítulo 2
De los genes a los ecosistemas
§. Principio de cooperaciónDe acuerdo con el marchante de arte Ambroise Vollard, que Cézanne pintara un retrato era una experiencia larga que sacaba de quicio. Cuando su retrato estaba a punto de concluir, Vollard cometió el error de comentar que había dos zonas que no le parecían terminadas:
§. Principio de la riqueza combinatoria
§. Nubes errantes
§. Principio de recurrencia
§. El origen de las especies
§. Especies y ecosistemas
§. Una receta para la evolución
Para alguien que no le ha visto pintar, es difícil imaginar hasta qué punto, algunos días, sus progresos eran lentos y dolorosos. En mi retrato hay dos pequeñas manchas en la mano donde el lienzo no quedó cubierto. Se lo señalé a Cézanne y me replicó: «… Podría encontrar el color justo para rellenar esas manchas blancas. Pero entienda, Monsieur Vollard, que si las cubro con uno al azar, me veré forzado a comenzar el cuadro de nuevo a partir de ese punto.»[12]Cézanne nunca enmendó estas dos manchas (se pueden ver todavía como motas blancas en la base del dedo corazón en la figura 5 [lámina 1]), por lo que Vollard se ahorró otra serie de sesiones. Cézanne era perfectamente consciente de la interconexión entre los colores de un lienzo. Cada mancha tendría que pintarse en una relación muy precisa con las otras, como si fuera la armonía del cuadro en conjunto la que emergiera de la interacción de los colores. Cualquier cambio que no fuera armonioso, como poner el color inadecuado en la mano, podría conjurar un desastre.

Figura 5. Retrato de Ambroise Vollard, Paul Cézanne, 1899. Véase la lámina 1.
En el capítulo anterior dimos por sentado que el mundo viviente está organizado en unidades espaciales denominadas individuos. Las variaciones entre estos individuos de una población dan pie a que intervenga la selección natural. Pero ¿por qué la vida está organizada de esta manera?
§. Principio de cooperación
Podríamos pensar que el principio de competencia conducirá a la selección natural hacia un egoísmo despiadado. Pero cuando nos enfrentamos a una competencia intensa, a menudo resulta ventajoso combinar las fuerzas y el trabajo, siempre y cuando esto produzca un beneficio mutuo[13]. Este tipo de interacción aflora sobre todo cuando los componentes están conectados o comparten el mismo espacio.Consideremos un organismo unicelular. Su ADN está formado por una larga secuencia de pares de bases concatenadas para formar los cromosomas. El ADN se recombina de una generación a otra gracias a que los cromosomas se rompen al azar y se vuelven a unir, con un cierto intercambio de piezas de ADN entre ellos (recombinación). Como estas roturas se producen solo en unas pocas posiciones durante la reproducción sexual, resulta muy poco probable que se recombinen las bases cercanas. Por esta misma razón, si estamos guardando una larga cola, en el caso de que esta se dividiera aleatoriamente en dos es muy poco probable que nos separemos de los vecinos más cercanos. Así que, en una molécula de ADN, las bases cercanas tienden a permanecer juntas de una generación a otra. Por consiguiente, si una base mutada en una posición del genoma favorece el éxito reproductor, y por ende su propia diseminación por la población, con toda probabilidad ayudará a que se diseminen junto a ella las bases que estaban cerca. De igual forma, una base proporcionará beneficio si alguna de las cercanas resulta que incrementa el éxito reproductor. Existe un incentivo mutuo de cooperación entre las bases cercanas porque su conexión física hace que se hereden juntas con mucha probabilidad. Con incentivar la cooperación no pretendo sugerir que una base piense en la otra, sino que lo que es beneficioso para una, en términos de éxito reproductor, probablemente también sea beneficioso para la otra. En tal situación, la selección natural favorece los casos en los que las bases cercanas colaboran con eficacia para asegurar el éxito reproductor. Por supuesto, las secuencias de las bases cercanas también podrían interaccionar negativamente para reducirlo, pero en ese caso es menos probable que la selección natural las fije y, en consecuencia, no llegarían a predominar. «Cooperación» se utiliza para referirse a los resultados colaborativos sin ningún propósito psicológico, del mismo modo que «competencia» se utilizó en el capítulo anterior para referirse a una situación donde hay muchas entidades y solo unas pocas logran ocupar un número limitado de sitios.
Una forma de cooperación entre las bases consiste en codificar una proteína. Las proteínas son moléculas grandes que en su mayoría sirven para incrementar la velocidad de determinadas reacciones químicas, tales como la degradación de la glucosa o la fijación del carbono. Cada proteína está formada por una cadena de subunidades moleculares denominadas aminoácidos. Hay hasta 20 tipos diferentes de aminoácidos en una proteína, o 20 letras en el alfabeto proteico. Las propiedades de una proteína dependen del orden preciso en el que se encadenan los aminoácidos, lo que a su vez viene determinado por un tramo de la secuencia del ADN (un gen) en el genoma. Cada tipo de proteína está codificado por su correspondiente gen. El genoma codifica muchos miles de proteínas, por lo que cada una de estas secuencias de ADN representa tan solo una fracción minúscula del ADN total.
Una proteína supone un esfuerzo cooperativo o colectivo porque está codificada por una cadena de bases cercanas en el ADN. Una única base no puede codificar una proteína, sino que se requiere una secuencia de ellas en un gen. Existe otra relación entre el gen y la proteína: ambos residen en la misma célula. La membrana que envuelve la célula impide que la proteína se aleje demasiado del gen y garantiza que el destino del gen y de la proteína queden entrelazados. Esto significa que si la proteína favorece la supervivencia y la reproducción de la célula, también incrementará el éxito reproductor del gen que la codifica; si la proteína tuviera total libertad para alejarse, no aportaría ningún beneficio directo al gen. Tenemos dos relaciones físicas muy estrechas: las bases dentro del mismo gen están ligadas, y las proteínas también se encuentran muy cerca por el confinamiento de la membrana celular. Esta intimidad espacial conecta el destino de estos componentes tan distintos; lo que favorece a uno en términos de éxito reproductor también favorece al otro. Por supuesto, esto también significa que si un componente reduce el éxito reproductor, también arrastrará al otro con él. Pero tales cambios no estarán favorecidos por la selección natural (la selección actuará en su contra) puesto que en las poblaciones tienden a establecerse las variaciones que incrementan la capacidad reproductora. Al estar en el mismo barco se alienta a que se ayuden mutuamente y a que se compartan los beneficios.
El razonamiento es el mismo cuando consideramos los demás genes de un organismo unicelular: todos conviven dentro de la misma célula, por lo que les sale a cuenta cooperar poniendo sus proteínas a trabajar juntas. Sin embargo, la intimidad entre los diferentes genes no es realmente tan fuerte como entre las bases del mismo gen porque las secuencias de ADN que están más distantes en el genoma tienen más probabilidad de barajarse y recombinarse a medida que transcurren las generaciones. Por lo tanto, es probable que una versión de un gen que surgió en un individuo se separe de sus vecinos originales y se junte con otra versión génica que surgió en otro individuo. La selección natural valora cada versión génica en relación con el abanico de genes con los que se topa en la población, y no únicamente con sus vecinos originales. La cooperación todavía es primordial porque cada gen necesita trabajar de forma eficaz con otros genes de una célula para favorecer su éxito reproductor, pero el impulso para la cooperación es menos intenso que para las bases cercanas.
En los organismos pluricelulares, la cooperación se extiende a todas las células en un individuo. A medida que un huevo fecundado crece y se divide, el ADN se copia con fidelidad, por lo que, con muy pocas excepciones, todas las células del adulto contendrán una copia de la misma secuencia de ADN. Se trata de una organización que favorece la colaboración: si las células de cada individuo cooperan, la diseminación de sus genes compartidos se favorecerá en la población mediante la selección natural. De nuevo, la proximidad física se revela de una importancia capital: si las células se alejaran en diferentes direcciones después de la fecundación, ya no serían capaces de ayudarse mutuamente con eficacia ni de compartir los beneficios que pudieran acumular.
Tendemos a dar por hecho el concepto de individuo. Parece obvio que los organismos existen como paquetes organizados en el espacio. Sin embargo, esta individualidad no sale de la nada, sino que es el resultado de dos factores subyacentes. El primero es que las entidades cercanas suelen interaccionar con más intensidad que las distantes. Se trata de una propiedad fundamental de la materia como consecuencia de la disminución de la intensidad de las fuerzas físicas, tal como la atracción o repulsión eléctricas, con la distancia. El segundo es que, en una situación competitiva, será rentable cooperar si los destinos están entrelazados. En el contexto de la selección natural se favorece la cooperación si las entidades se encuentran en el mismo barco, con lo que se facilita la ayuda mutua y se pueden compartir los beneficios sobre la reproducción. Esto conduce a que colaboren los componentes que mantienen una conexión espacial cercana, ya sean bases en el ADN, genes, proteínas o células. La proximidad ayuda a vincular las actividades y los destinos. Los primeros organismos vivos pudieron haber establecido colaboraciones relativamente toscas, pero durante miles de millones de años de evolución se modificaron por selección natural, lo que dio lugar a los individuos que vemos hoy en día.
Dado que las formas de vida se organizan en unidades cooperativas, solo seremos capaces de juzgar la importancia de cualquier cambio en un componente si tenemos en cuenta su manera de interaccionar con los otros. El efecto de un cambio de base depende de la secuencia de ADN que la rodea. La base por sí sola no dice prácticamente nada, al igual que una única letra de una palabra significa muy poco cuando está aislada. Lo mismo vale para los aminoácidos: la consecuencia de cambiar un aminoácido depende de su interacción con todos los demás de la proteína. De igual forma, el efecto debido al cambio de una proteína depende de sus interacciones con el enorme conjunto de proteínas del individuo. El concepto de que la selección natural actúa sobre una base o sobre un gen aislado no tiene mucho sentido, porque las bases y los genes solo ejercen su efecto al interaccionar con otros componentes, interacciones que pueden suceder a muchos niveles: las bases adyacentes en un gen, los diferentes componentes dentro de una célula, los grupos de células en un tejido, o los diferentes órganos de un individuo. Sería equiparable a las interacciones en muchos niveles entre los colores de las pinturas de Cézanne. Hemos visto que la cooperación emergió de resultas de la selección natural. Pero la cooperación también se puede ver como un ingrediente de la selección natural, porque sin cooperación, la selección natural no tendría individuos ni genes sobre los que actuar. La competencia y la cooperación se alimentan mutuamente. La competencia conduce a unidades espaciales cooperativas, que a su vez proporcionan los conjuntos que impulsan más competencia. Esta realimentación continua entre la competencia y la cooperación es una característica fundamental de la evolución por selección natural, incluso aunque a veces no nos demos cuenta. La selección natural se suele describir como algo más relacionado con la competencia que con la cooperación; es el camino que también yo seguí cuando la presenté en el capítulo anterior. A pesar de todo, tales descripciones dan por sentado la existencia de unidades cooperativas, como los individuos o los genes. Si nos limitamos a fundamentarla en la competencia, estaremos ignorando el bucle de realimentación que une la competencia con la cooperación. Por naturaleza, los lazos de realimentación no tienen un punto de partida claro; dar primacía a la competencia introduce una ruptura artificial en lo que es realmente un ciclo de interdependencia. La cooperación es un principio de cambio evolutivo que resulta tan fundamental como la competencia.
§. Principio de la riqueza combinatoria
En una de las semblanzas de sus pacientes de neurología, Oliver Sacks describe el caso del señor I, un pintor que padeció acromatopsia (ceguera a los colores) a los 65 años después de un accidente de coche. Antes era un artista consumado con una apreciación sutil del color. Después del accidente, el zumo de tomate parecía negro y su perro pardo parecía gris. «A medida que transcurrían los meses echó de menos especialmente los colores brillantes de la primavera: siempre le habían gustado las flores, pero ahora solo conseguía distinguirlas por su forma u olor. Los arrendajos azules ya no eran brillantes y su azul, curiosamente, se veía ahora como un gris pálido[14] ». El señor I experimentaba un sentimiento de pérdida insufrible y padeció una depresión que casi le lleva al suicidio a las pocas semanas del accidente.Damos por sentada la riqueza del mundo y solo apreciamos totalmente su diversidad cuando desaparece una característica determinada. Por supuesto, el color es solo un atributo de los muchos que apreciamos. Sin llegar a ver los colores, el señor I podía seguir percibiendo la llegada de la primavera, la forma y olor de las flores y el vuelo de un pájaro. Al hacerlo, estaba captando la complejidad del mundo en una serie de escalas diferentes, de la órbita de la Tierra alrededor del Sol hasta la forma o movimiento de los objetos cercanos, o hasta las sustancias químicas que emite una flor.
Pero supongamos que la Tierra fuera solo un terrón monótono y aburrido de materia sin ninguna variación digna de mención. Independientemente de los sentidos que poseamos, el mundo parecería sin interés porque no habría nada que nos embriagara. Las consecuencias, sin embargo, irían mucho más allá: ni siquiera estaríamos aquí para contemplar un mundo tan tedioso porque la complejidad de los organismos vivos (lo que nos incluye a nosotros mismos) está relacionada con la complejidad del mundo en el que viven. La selección natural solo puede desembocar en adaptaciones si hay peculiaridades ambientales a las que adaptarse. Nuestra capacidad para contemplar lo que nos rodea solo ha evolucionado porque hay un entorno que merece la pena contemplar. Si el mundo fuera realmente aburrido, también nosotros seríamos aburridos.
Por supuesto, nuestro mundo no es tan aburrido: la Tierra contiene muchos elementos atómicos diferentes que son capaces de combinarse entre sí según las leyes de la física y de la química. Aunque haya tan solo unos cien tipos diferentes de átomos, la cantidad de formas en las que consiguen combinarse es mucho más numerosa. El hidrógeno y el oxígeno se combinan para formar moléculas de agua (H2O). El sodio y el cloro se asocian para formar la sal. Además, estas moléculas consiguen combinarse entre sí de varias maneras: las moléculas del agua se juntan para formar el hielo, nieve, ríos, vapor o nubes, y la sal se combina con el agua para dar lugar a una solución de cloruro de sodio. Se puede generar un amplio abanico de ordenamientos simplemente por la unión de unos pocos elementos de diferentes maneras, en lo que denomino el principio de la riqueza combinatoria, que es lo que convierte nuestro mundo en un lugar interesante y rico.
Por consiguiente, los organismos afrontan numerosos retos para lidiar con el mundo que los rodea, retos que van satisfaciendo gracias a la abundancia interna de disposiciones moleculares. Esta abundancia de posibilidades surge de nuevo por el principio de riqueza combinatoria. Para ver cómo, consideremos una situación más bien irreal en la que podríamos variar a voluntad la longitud del ADN de un organismo, mientras que el organismo sigue pudiéndose reproducir. Supongamos que tallamos el ADN al máximo para dejarle una longitud de un solo par de bases, con lo que solo podrían existir dos tipos de genomas: uno con una A en una hebra de ADN y T en la opuesta, o bien una G en una cadena y una C en la opuesta. No importa lo interesante que sea el mundo, tales organismos solo podrían elegir entre dos posibles configuraciones de ADN.
Si permitimos que los genomas tengan dos pares de bases, tendría más posibilidades. Por ejemplo, podríamos tener organismos con AA, AG o AC en una hebra del genoma. Podemos representar todas las posibilidades con un diagrama en el cual la base de la primera posición variara en el eje horizontal y la segunda base variara a lo largo del eje vertical (figura 6). El cuadrado resultante indica 16 organismos posibles (4 × 4, o 4 2).Sin embargo, dado que cada una de estas secuencias de bases representa solo una de las dos hebras del ADN, algunas de ellas representan la misma secuencia de genoma (AA, por ejemplo, es lo mismo que TT visto desde la otra hebra), lo que significa que tenemos que dividir el número total de combinaciones entre dos, lo que nos dará ocho genomas diferentes (42/2 = 8). Esto contrasta con las dos posibilidades para los genomas que miden un solo par de bases. Con solo añadir el mismo tipo de unidad, otra base, hemos multiplicado las posibilidades por cuatro.
Figura 6. Secuencias posibles para una hebra de una molécula de ADN de dos pares de bases de largo.
Figura 7. Secuencias posibles para una hebra de una molécula de ADN de tres pares de base de largo. Solo se muestran tres superficies externas del cubo, pero hay más posibilidades en el interior.
¿Qué ocurre con un genoma de cien pares de bases? La representación de todas las posibilidades requeriría un hipercubo con 100 dimensiones, y no somos capaces de visualizar tal objeto (incluso Einstein se esforzó para pensar en cuatro dimensiones, no digamos en cien). La presencia de muchas dimensiones es una manera cómoda de decir que podemos variar muchas peculiaridades de forma independiente las unas de las otras. Debido al principio de la riqueza combinatoria, cuantas más dimensiones tengamos, más inmensamente mayor será el número de posibilidades. Para un hipercubo de 100 dimensiones con cuatro bases en cada eje, hay 4100/2genomas diferentes, que son unos 1060 (un 1 con 60 ceros detrás). Se trata de un número astronómicamente grande, mucho mayor que el número de organismos que han habitado en la Tierra[15]. Basta con repetir la misma unidad cien veces y dejar que cada unidad varíe entre cuatro posibles estados para que creemos un espacio enorme de posibilidades. Estamos ante la manifestación del principio de la riqueza combinatoria, en la que los componentes variables son los módulos moleculares (en el ADN son las bases nitrogenadas).
Al igual que el abrumador abanico de posibilidades de variación del ADN, a las proteínas les corresponde otro igualmente enorme. Dada una proteína de 100 aminoácidos, el abanico de posibilidades también se puede representar con un hipercubo de 100 dimensiones, pero en este caso en cada eje hay 20 divisiones en vez de cuatro (que corresponden a los 20 aminoácidos diferentes). Esto arroja 20100(aproximadamente 10130) posibilidades, mucho más que el número de átomos del universo observable, que es tan solo de 1078.De nuevo, el principio de la riqueza combinatoria crea este espacio enorme de proteínas posibles.
Estos hiperespacios de 100 dimensiones no son más que la punta del iceberg: el genoma humano contiene unos 25.000 genes que codifican proteínas, cada uno con varios miles de bases de longitud. Si consideramos el intervalo de posibilidades con 25.000 dimensiones de genes, cada una de las cuales con su enorme hipercubo de posibilidades, nos enfrentamos a un hiper hipercubo colosal. Lo mismo ocurre con las 25.000 proteínas que están codificadas por estos genes. El intervalo de combinaciones de aminoácidos y proteínas es verdaderamente abrumador.
Cuando se secuenció por primera vez el genoma humano, muchos científicos esperaban que contuviera más genes que el de otros organismos, simplemente porque parecemos mucho más complejos que cualquier otra especie. Las estimaciones iniciales colocaban el número de genes en los humanos alrededor de 70.000, más que para los otros organismos. Pero a medida que conocíamos mejor el genoma, este número no hacía más que disminuir. Las estimaciones más recientes colocan el número de genes que codifican proteínas en el genoma humano en torno a 25.000, aproximadamente el mismo número de genes que en un hierbajo llamado Arabidopsis. En un principio esto puede resultar sorprendente porque los humanos parecemos mucho más complicados que los hierbajos, pero incluso 25.000 genes contienen muchísimas más posibilidades de las que podemos concebir. La vida es interesante debido a la enorme riqueza combinatoria de los organismos y de sus entornos.
§. Nubes errantes
Para hacernos una idea de lo que significa para la evolución tener a su disposición muchas posibilidades genéticas, conviene imaginarse el cambio evolutivo. Dado que no es fácil concebir grandes hiperespacios, una alternativa consiste en imaginar un espacio tridimensional inmenso. Pensemos en todas las posibilidades genéticas como si fueran un espacio enorme en el que cada punto corresponde a una secuencia concreta del genoma. Entonces, el genoma de cualquier individuo correspondería a una posición en este inmenso espacio genético de posibilidades. La secuencia del genoma de uno de nosotros, por ejemplo, se encontraría en una posición concreta del espacio genético. Cada persona tiene una secuencia genómica ligeramente diferente debido a las mutaciones y a que los cromosomas se están continuamente barajando y recombinando. Por consiguiente, la población humana en conjunto correspondería a un conglomerado de puntos, o a una nube, en el espacio genético. Esta nube sería muy pequeña en relación con todo el espacio porque cada especie solo representa una fracción insignificante de las posibilidades de variabilidad genética. Como en el planeta viven muchas especies, habría muchas nubes como esta, y cada una ocuparía una parte diminuta del espacio genético.Ahora podemos pensar que la evolución no es más que desplazamientos de la posición de las nubes de población, lo que corresponde a cambios de la composición genética de las especies. Durante la evolución, cada nube de población explora de hecho determinadas regiones del espacio genético, avanzando en varias direcciones. Como hay muchas especies, hay muchas nubes en movimiento por el espacio genético en todo momento. A pesar de todo, al ser tan inmenso, la vida en la Tierra solo ha explorado una fracción diminuta.
Para hacerse una idea más concreta de lo que esto significa, miremos cómo podría evolucionar una única proteína. Las plantas son capaces de fijar el dióxido de carbono de la atmósfera gracias a una proteína llamada ribulosa-1,5-bisfosfato carboxilasa, o Rubisco de forma abreviada. Esta proteína se une a moléculas de dióxido de carbono de la atmósfera y las anima a fijarse a otras moléculas de carbono. A primera vista, la Rubisco parece que fija el dióxido de carbono con eficacia al ser capaz de fijar cerca de tres moléculas de carbono cada segundo, lo que da la impresión de rapidez. Pero la realidad es que es una proteína más bien lenta, ya que otras completan cientos o miles de reacciones por segundo. Por lo tanto, la Rubisco es más bien perezosa. La lentitud no es su único defecto, porque también comete errores con frecuencia, como fijar oxígeno en vez de dióxido de carbono. La fijación del oxígeno es un derroche para la planta porque acaba conduciendo a la liberación de dióxido de carbono, lo contrario a la fijación del carbono, de manera que las plantas pierden hasta un tercio del carbono que fijan debido a este error molecular. El problema es que la forma de la Rubisco no es lo suficientemente específica para discriminar siempre entre el dióxido de carbono y las moléculas de oxígeno.
El problema se vuelve particularmente acuciante en las plantas que viven en ambientes secos y calurosos, en los que los estomas de las hojas se abren lo menos posible para impedir la pérdida de agua. Los estomas cerrados hacen caer la concentración de dióxido de carbono en la hoja porque el carbono se fija mucho más rápido de lo que se reemplaza desde la atmósfera. Entonces la Rubisco comete más errores: hay menos moléculas de dióxido de carbono que de oxígeno, por lo que este último se unirá con más frecuencia que el primero. Varias especies vegetales han evolucionado para solucionar este problema, de manera que la Rubisco solo se sintetice en unas células especializadas de las hojas, mientras que las otras bombean la poca cantidad de dióxido de carbono hacia las células especiales, lo que hace aumentar su concentración de forma local. De esta manera, incluso aunque la concentración global de dióxido de carbono de la hoja sea baja, será elevada en las células especializadas con la proteína Rubisco, y se reducirá así la probabilidad de error. Así pues, la Rubisco está lejos de ser óptima. Aunque fije el carbono de forma razonable, quizá exista en algún lugar del espacio de proteínas posibles otra forma de la proteína que fijaría el dióxido de carbono con más rapidez y eficacia. La evolución nunca ha llevado a esta versión de la proteína porque solo se ha explorado una diminuta fracción de las posibilidades. Pero aunque la Rubisco no sea la óptima, la selección natural ha dado pie a otras maneras de solventar estas limitaciones. Una consiste en producir una gran cantidad de la proteína para que haya suficiente cantidad y se fije el carbono necesario. De hecho, las plantas producen tanta Rubisco que se piensa que es la proteína más abundante del planeta. El segundo método, como hemos visto, consiste en bombear el dióxido de carbono a las células especializadas para elevar su concentración local. Por lo tanto, en vez de bloquearse por las limitaciones de la Rubisco, la evolución ha encontrado otras vías y estratagemas con las que solventar el problema.
Podríamos considerar que las soluciones basadas en producir más proteína o en bombear el dióxido de carbono dentro de la hoja son chapuzas abominables que ponen de manifiesto la incompetencia de la selección natural para encontrar una proteína óptima. O del mismo modo podríamos contemplar las soluciones como hermosas adaptaciones que funcionan dentro de las restricciones del sistema. De cualquier modo, lo que importa es que los problemas se pueden soslayar de todas las maneras posibles porque el espacio genético es inmenso y aún le faltan por explorar muchas posibilidades.
Esta flexibilidad significa que los problemas a menudo se solucionan de diversas maneras a medida que avanza la evolución. Consideremos la evolución del vuelo. Las plumas fueron el rasgo crucial que permitió que el vuelo evolucionara en las aves. Por lo tanto, podría parecer que la falta de plumas bloquearía la evolución del vuelo en otros grupos, como los mamíferos. Aun así, el vuelo ha evolucionado en los mamíferos, como los murciélagos, con una solución diferente para el mismo problema: en vez de plumas, sus alas están formadas por alerones extendidos de piel, estirados entre sus dedos largos y delgados. De igual forma podemos comparar las diferentes formas de trinar: las aves trinan por la vibración de membranas en las vías aéreas, mientras que los grillos frotan un ala contra la otra. Las diferentes opciones genéticas posibilitan diferentes soluciones.
Un espacio genético inmenso de posibilidades tiene dos consecuencias. La primera es que las nubes de población errantes solo han visitado una pequeñísima parte de este espacio. La segunda es que la alta dimensionalidad del espacio significa que hay muchas direcciones en las que se pueden mover las nubes, lo que proporciona una flexibilidad enorme. Lo que la selección natural pierde al no ser capaz de cubrir todas las posibilidades lo gana en el número de direcciones que puede tomar.
Las creaciones artísticas humanas se pueden contemplar desde una perspectiva parecida. Una de las limitaciones de un cuadro es que no puede mostrar el movimiento directamente. Sin embargo, los artistas han probado distintas estratagemas para transmitir movimiento a pesar de las cortapisas. Un buen ejemplo es el cuadro de Miguel Ángel La creación de Adán (figura 8), en el techo de la Capilla Sixtina. El movimiento hacia la izquierda de Dios se transmite mediante el ropaje ondulante y el cabello azotado por el viento, además del brazo extendido y la brecha sugerente en la punta del dedo que está pidiendo que la cierren.
El impresionista Claude Monet utilizó una estrategia diferente para transmitir el movimiento más de 350 años después. En La calle Montorgueil, Monet pintó numerosas banderas de un modo impreciso y embrollado (figura 9, lámina 3). Mediante esta falta de definición, consiguió un sentido notable de movimiento y vitalidad. Nuestros ojos parecen incapaces de detenerse en nada y por eso las banderas parecen agitarse ante nosotros. Después de Miguel Ángel podría pensarse que un cuadro solo conseguiría representar más movimiento retorciendo más el cuerpo, o encrespando las ropas todavía más. De hecho, muchos seguidores de Miguel Ángel intentaron capturar el movimiento precisamente de este modo, con contorsiones corporales cada vez más elaboradas. Gracias a que siguió una estrategia diferente, Monet transmitió el movimiento de un modo totalmente nuevo.

Figura 8. La creación de Adán, Miguel Ángel Buonarroti, 1510.
§. Principio de recurrencia
Ya hemos tratado seis principios de la evolución. En el capítulo anterior hemos visto que cuatro principios (variabilidad de la población, persistencia, refuerzo y competencia) interaccionan para proporcionar los fundamentos de la selección natural. En este capítulo han aflorado dos principios más: uno es el principio de cooperación (la proximidad permite que los destinos se entrelacen, por lo que la selección natural conduce a la formación de unidades colaborativas, como genes, proteínas, células e individuos; estas unidades cooperativas son tanto un resultado como un ingrediente de la selección natural) y el otro principio es la riqueza combinatoria que conduce a un mundo de diversidades y a un inmenso espacio de posibilidades genéticas.
Figura 9. La calle Montorgueil, París, celebración del 30 de junio de 1878, Claude Monet, 1878. Véase la lámina 3.
La respuesta implica el modo en el que la selección natural se realimenta a sí misma. Podría parecer que una vez que el organismo se ha adaptado mejor a su medio, la selección natural sería menos intensa. Pero no es así. Para ver los motivos, imaginemos que estamos compitiendo en una carrera automovilística y que inventamos un artilugio que hace que nuestro coche vaya más rápido que los otros. Al principio tendremos ventaja y ganaremos, pero una vez que todo el mundo conozca el artilugio y lo incorporen en todos los coches, perderemos la ventaja porque todos los automóviles ahora irán más rápido. Lo que fue al comienzo una ventaja, ahora es simplemente un rasgo estándar de todos los coches, un nuevo contexto en el cual se pueden realizar otras mejoras. En vez de reducir la competencia, la difusión del artilugio ha mejorado simplemente el rendimiento de cada uno, y ha empujado a los inventores a que desarrollen artilugios aún mejores para que sus coches ganen la carrera.
De igual forma, una vez que una adaptación consigue fijarse por selección natural, la compartirán todos los individuos de la población. La competencia será incluso más feroz que antes porque ahora todos los individuos se reproducen mejor y compiten por los escasos recursos. La selección natural no se debilita a medida que favorece la diseminación de las adaptaciones, sino que se estimula a medida que cada adaptación desplaza el contexto y hace que la competencia sea cada vez más intensa. Nuestras nubes de población imaginarias se impulsan a sí mismas por el espacio genético, de manera que cada nueva adaptación crea las condiciones que ayudan a surgir la siguiente. Del mismo modo que una carrera automovilística no persigue alcanzar una velocidad determinada, sino ir más rápido que los demás coches, la selección natural no pretende que se alcance un determinado nivel de adaptación, sino que funciona de un modo relativo: favorece continuamente las versiones génicas que sirven para una reproducción más eficaz con respecto al resto de la población. No hay «tiempo muerto» para la selección natural porque la competencia nunca se para, sino que se intensifica con cada adaptación que se lleva a cabo.
A esta repetición implacable de un proceso, siempre incitada por lo que había antes, la denomino el principio de recurrencia. Sin recurrencia, los organismos representarían tan solo un burdo intento de capturar una única variable ambiental, el resultado de una única etapa de la selección natural. En cambio, tenemos un proceso continuo de adaptación que se fundamenta en la adaptación previa, impulsado por los estándares siempre cambiantes que va creando. En consecuencia, nuestras nubes de población raramente permanecen quietas, sino que están en movimiento continuo a medida que los nuevos contextos crean más desafíos[16].
La evolución se ve propulsada continuamente por el pasado, que siempre modifica su propio contexto y crea las condiciones para las nuevas etapas. Esta dependencia del pasado hace que sea un proceso muy constreñido por la historia. La proteína Rubisco, por ejemplo, comenzó su evolución hace miles de millones de años cuando había poco oxígeno en la atmósfera, por lo que su incapacidad para discriminar entre las moléculas de oxígeno y de dióxido de carbono no tendría mucha importancia para las plantas. Incorporar oxígeno en vez de dióxido de carbono acabó convirtiéndose en un problema importante más tarde, cuando la fotosíntesis vegetal incrementó la concentración de oxígeno en la atmósfera. Pero para entonces podría resultar ya difícil modificar la estructura de la Rubisco para solucionar este problema debido a que la estructura ya presentaba determinadas características restrictivas. De igual forma, la evolución del vuelo de las aves y de los murciélagos estaba constreñida por la distribución particular de los huesos en las patas delanteras de sus ancestros. En el caso de las aves, las alas corresponden a brazos completos, mientras que en los murciélagos corresponden al estiramiento de la mano. No obstante, la distribución básica de los huesos en las patas delanteras se conserva en ambos casos y procede de la de sus ancestros. El precio de aprovechar siempre lo que ya existe son las limitaciones que provocan los primeros sistemas, limitaciones que no deben impedir que ocurran otros cambios porque, tal y como hemos visto, la riqueza combinatoria presenta muchas opciones evolutivas. Pero las limitaciones históricas a menudo dan lugar a trayectorias evolutivas muy enrevesadas e idiosincrásicas.
Se puede aplicar lo mismo a nuestra analogía artística. Las innovaciones de Monet dependieron de reacciones a una serie de logros anteriores en la pintura, entre ellos, los avances de Miguel Ángel y otros durante el Renacimiento. Si Monet hubiese nacido 400 años antes, no podría haber pintado cuadros de estilo impresionista. De igual forma, los descubrimientos renacentistas dependieron de los desarrollos artísticos del periodo bizantino y de la antigüedad clásica. Incluso aunque el arte se base en los logros anteriores, la historia del arte no es un camino recto. En algunos aspectos, el estilo impresionista puede parecer más tosco que el realismo del Renacimiento, y la pintura bizantina con sus figuras alargadas y aplastadas puede parecer más primitiva que la pintura de la antigüedad clásica que la precedió. Al igual que con la evolución biológica, el arte sigue caminos tortuosos porque está en continua construcción y reacción a lo que había antes, con lo que desplaza su propio contexto.
§. El origen de las especies
Hasta ahora nos hemos fijado en la forma de funcionar de la recurrencia dentro de una única población. Al fundamentarse constantemente en lo que había antes, en donde un contexto condiciona la situación para el siguiente, nuestra nube de población se mueve por el espacio genético para explorar diferentes regiones. Pero la evolución no afecta tan solo a una única población, sino que al haber muchas especies, cada una de las cuales se puede considerar una población de individuos que se cruzan entre sí, habrá muchísimas nubes de población, o de especies, explorando las distintas direcciones. Además, estas especies pueden interaccionar con otras, como vimos con las manzanas y los osos, y las interacciones proporcionan un contexto incluso más amplio para la evolución al abarcar ecosistemas completos. ¿Cómo surge este contexto más amplio?
Para responder esta cuestión, primero necesitamos conocer lo que mantiene unida a una población. ¿Qué impide que una nube de población se disperse por el inmenso espacio genético que la rodea? La respuesta tiene que ver con el sexo. Si un individuo es muy diferente del resto de su población, podría ser incapaz de aparearse de manera eficaz. E incluso si lo hiciera, la descendencia podría ser como un batiburrillo de rasgos diferentes que no tendrían fácil su supervivencia. Por consiguiente, el cruce continuo de individuos, combinado con la selección natural, garantiza que la variabilidad de una población esté confinada dentro de determinados límites. Este confinamiento mantiene en el espacio genético la cercanía de los miembros de la misma población reproductora. El sexo frecuente proporciona la cohesión, el pegamento genético que mantiene unida a la población y la define como una unidad o especie.
Pero esto conlleva un problema, el mismo que le provocó noches de insomnio a Darwin: si la reproducción sexual mantiene las especies juntas, ¿por qué tenemos tantas poblaciones diferentes en la Tierra, millones de especies diferentes? Si imaginamos que todas descienden de un ancestro común, entonces en algún momento esta especie ancestral debió dividirse en dos o más. Es de suponer que la sucesión de rondas de división como esta hayan dado lugar a las muchas especies de hoy en día. Pero ¿cómo puede ser que el pegamento sexual que mantiene unidas a las especies acabe desapareciendo, de modo que una especie se convierte en dos? Es difícil ver que la selección natural pueda bastarse por sí sola para impulsar este proceso de especiación. La selección natural favorece un incremento de la capacidad reproductora, a pesar de que un rasgo clave de dos especies es su incapacidad para procrear entre sí. Incluso si se aparearan con éxito, dos especies diferentes suelen producir una descendencia estéril, como cuando un burro y una yegua se aparean para producir una mula estéril. No es fácil ver cómo se podría seleccionar la infertilidad, que es lo contrario al éxito reproductor. Como he argumentado que la reproducción sexual mantiene unidas nuestras nubes de población, es difícil ver que puedan acabar dividiéndose, escindiéndose en dos.
Nos topamos con el mismo problema en los idiomas. Una de las características más raras del ser humano es que conversa en muchos idiomas diferentes. En aras de la comunicación, tendría más sentido para todo el mundo hablar el mismo idioma. La Biblia se detiene a explicar largo y tendido la razón por la que el Creador tuvo que imponer la diversidad lingüística. Según el libro del Génesis, al principio todo el mundo hablaba el mismo idioma y las diferentes lenguas surgieron más tarde como un castigo impuesto por Dios a los habitantes de Babel. Ocurrió cuando estas personas decidieron celebrar sus logros construyendo juntos una gran torre (figura 10). Tal acto de vanidad trajo la desaprobación de Dios, que castigó a los habitantes de Babel haciéndoles hablar diferentes idiomas, lo que destruyó su capacidad colaboradora y favoreció su dispersión, dando lugar a los muchos idiomas que se hablan hoy en día. Se trata de una extraña historia diseñada para explicar una elección extraña: si tuviéramos que crear humanos, tendría más sentido hacer que todos hablasen un mismo idioma, no varios diferentes.
Podemos plantear una explicación más prosaica del surgimiento de la diversidad lingüística. Consideremos los idiomas europeos, como el inglés, el francés o el italiano. Son tan diferentes que los hablantes de uno no pueden comprender fácilmente a los del otro. Aun así, todos ellos comparten raíces comunes: se remontan a una lengua ancestral común, que se cree que hablaban las primeras tribus fundadoras de Europa[17]. Si imagináramos todos los idiomas concebibles como un enorme espacio de posibilidades, entonces es como si una nube ancestral en este espacio lingüístico hubiera acabado dando lugar poco a poco a muchas nubes diferentes que constituirían las diferentes lenguas de hoy en día. A lo largo de este proceso, cada nube lingüística ha permanecido coherente (los individuos que comparten el mismo idioma siempre han sido capaces de conversar entre sí), y a pesar de ello, con el tiempo, se han separado tanto que se ha vuelto muy difícil cualquier conversación entre los individuos de nubes diferentes.

Figura 10. La Torre de Babel, Pieter Brueghel el Viejo, 1563.
Cada cambio por sí solo no altera la comprensión, pero con la acumulación de muchos, los idiomas pueden evolucionar y volverse muy diferentes unos de otros, tan diferentes que los hablantes de un idioma ya no conseguirán comprender a los del otro. En nuestro espacio lingüístico imaginario, cada idioma forma una nube que se mantiene pequeña porque solo hay variaciones lingüísticas menores entre las personas que suelen conversar entre sí. Pero cuando surgen barreras para la conversación interpersonal, con mucha frecuencia porque están distantes desde el punto de vista geográfico, las nubes lingüísticas pueden separarse y alejarse con el tiempo. Por supuesto, esta explicación de la divergencia lingüística está muy simplificada, pero ilustra la división de las nubes incluso si no hay nada que las incite activamente a separarse.
Puede surgir una divergencia equivalente en el caso de la evolución biológica. Las barreras físicas abundan en la Tierra porque la materia se agrega en escalas muy diferentes, y genera un mundo heterogéneo lleno de fronteras: los ríos, los mares o las montañas dividirán las regiones, y a menor escala, los estanques estarían divididos por franjas de tierra. Por lo tanto, las poblaciones verán restringida su capacidad de movimiento a una escala u otra. De vez en cuando, unos pocos individuos de una población consiguen superar una barrera e invadir una nueva localidad, al nadar a una isla o al viajar de un estanque a otro, y con ello crearán una nueva colonia. El resultado son dos poblaciones separadas en el espacio. Como la reproducción sexual es un acto cooperativo que depende del contacto físico (la unión de las células sexuales de cada progenitor), el material genético se intercambiará poco o nada entre estas dos poblaciones.
En ausencia del contacto, nuestras dos nubes de población comenzarían a apartarse hacia regiones diferentes del espacio genético. Esta divergencia puede verse impulsada en parte por las diferencias ambientales entre las dos localizaciones, que conducirán a adaptaciones diferentes por selección natural. Pero incluso sin diferencias ambientales importantes, podríamos esperar que las nubes se separaran, quizá por los compromisos: a menudo se es mejor en un aspecto a expensas de ser peor en otro. Tener pezuñas, por ejemplo, puede significar que eres más bueno corriendo por el suelo, pero te puede hacer peor para trepar a los árboles; se obtiene una ventaja a costa de una desventaja. Para algunos cambios evolutivos, los beneficios obtenidos de adaptarse en un cierto aspecto pueden compensarse más o menos con las pérdidas en otro. La ventaja de ser capaz de correr más rápido puede quedar contrarrestada por la desventaja de no ser capaz de trepar a los árboles. En tales casos, un cambio evolutivo quizá no produzca ninguna diferencia importante en la capacidad global de un organismo para sobrevivir y reproducirse, incluso aunque modifique su funcionamiento. En términos evolutivos, el cambio es casi neutro, equivalente en nuestra analogía lingüística a sustituir una manera de decir manzana por otra igualmente buena. Cuantas más dimensiones tenga nuestro sistema, más probable es que tales compromisos surjan, porque habrá más opciones que explorar[18]. Dadas las muchas dimensiones del espacio genético y el entorno, cabe esperar que haya muchas trayectorias de esta clase por las que puedan moverse nuestras nubes de población. Los compromisos son habituales porque la riqueza combinatoria garantiza que la vida pueda tomar muchas direcciones equivalentes.
Si una población queda dividida por una barrera física, como una cadena montañosa, resulta probable que las poblaciones separadas comiencen a alejarse la una de la otra en el espacio genético y a operar de diferentes maneras, del mismo modo que los idiomas separados divergen con el tiempo. Al apartarse de esta forma, las poblaciones también podrían dejar de ser compatibles desde el punto de vista sexual, al igual que la comunicación entre los que hablan lenguajes divergentes se hace cada vez más difícil. La incompatibilidad sexual no está empujada por la selección natural, sino que es una consecuencia indirecta de la barrera reproductora entre las nubes. Para decirlo de otro modo, al igual que la comunicación, la compatibilidad sexual entre los individuos de una población se tiene que mantener activa con intercambios regulares; una vez que se detienen estos intercambios, las incompatibilidades comenzarán a acumularse. He dado un ejemplo de cómo la separación de las nubes (la división de una especie en dos) puede tener lugar después del aislamiento geográfico, pero no se debe pensar que no existen más mecanismos: hay muchos otros escenarios posibles con varios grados y formas de separación.
§. Especies y ecosistemas
Hemos visto que una especie puede dividirse en dos después de que se separen por una barrera física. Pero ¿cómo ha conseguido la evolución conducir a la formación de varias especies en el mismo lugar? Esto implica que muchas nubes del espacio genético coexisten en la misma región de espacio físico. Para responder esta pregunta, miremos los posibles resultados cuando se restablece algún contacto entre poblaciones después de una separación temporal.Supongamos que unos pocos individuos de una de nuestras poblaciones cruzan la barrera separadora y llega al área habitada por otra población. Si los recién llegados aún pueden aparearse con los de la población que se encuentran, se podrían integrar totalmente en ella poco a poco a lo largo de varias generaciones. La nube genética de los recién llegados se absorbe en la de la población hospedadora.
Otra posibilidad tiene lugar cuando las poblaciones han divergido tanto que ya no consiguen aparearse, en cuyo caso los recién llegados formarían una cohorte distinta que coexiste con la población más grande. Si la nueva cohorte vive de un modo muy similar a sus vecinos y se mezcla libremente con ellos, seguramente sobrevendrá la competencia por los mismos y escasos recursos. Esto podría finalmente conducir a la extinción de una de las poblaciones debido bien a que es menos capaz de competir, o bien a las leyes del azar (si ambas poblaciones compiten igualmente bien). Sin embargo, si los recién llegados funcionan de modo diferente a sus vecinos, podrían explotar el entorno de diferentes maneras y, por lo tanto, ocupar un nicho ligeramente diferente. Es probable que estén disponibles muchos nichos debido a la riqueza combinatoria de los medios físicos y de los organismos que están en ellos. Al ocupar nichos ligeramente diferentes, las poblaciones podrían coexistir de forma más estable porque cada una podría mantenerse con recursos que le son hasta cierto punto peculiares. Como resulta beneficioso que no compitan por los mismos recursos, cabría esperar que la selección natural dentro de cada población incrementara la especialización, con lo que las poblaciones seguirían divergiendo con el tiempo. En otras palabras, las dos nubes genéticas se separarían incluso más, como si se repeliesen entre sí. El resultado es que las dos nubes de población se distancian cada vez más en el espacio genético, incluso aunque ocupen la misma región del espacio físico.
Hemos acabado con un pequeño ecosistema que comprende dos especies, donde cada una explota un nicho diferente. Debo insistir en que estos nichos no son rasgos absolutos del medio, sino que surgen por las interacciones entre las dos poblaciones. Si solo hubiera una población, el concepto de nicho sería innecesario, ya que los nichos solo hay que definirlos cuando consideramos poblaciones que interaccionan y que se especializan. Un nicho es un concepto relacional, una peculiaridad que evoluciona junto con la especie, en vez de existir de forma independiente.
Con poblaciones que continuamente se dividen, separan y vuelven a encontrarse, pueden evolucionar múltiples especies, cada una de las cuales ocupará un nicho distintivo en un ecosistema. El resultado es muchos organismos que viven juntos y cada uno captura aspectos concretos de su entorno. Por supuesto, existe un límite para esta división de nubes porque los ecosistemas ocupan regiones finitas y no pueden mantener un número infinito de especies. Al igual que se forman nuevas especies, otras se acaban extinguiendo. Los ecosistemas de nuestro mundo son el resultado de este equilibrio entre la producción y la extinción de las especies[19]. Debido a la actividad humana, la tasa de extinción de las especies es actualmente mucho más elevada que la tasa a la que se originan, por lo que su número está disminuyendo rápidamente.
Como los ecosistemas maduros contienen muchas especies, hay muchas formas complejas en que nuestras nubes de población pueden interactuar, dividirse y apartarse. Los compromisos pueden surgir ahora mediante interacciones entre las especies. Los azúcares que hacen que una manzana sea más dulce para un oso también pueden hacerla más atractiva para los hongos y las bacterias: un logro en la dispersión de las semillas se ve comprometido al incrementarse la susceptibilidad a enfermedades. La selección natural sobre la población de manzanos actuaría incrementándole la resistencia a las enfermedades, pero al mismo tiempo la selección natural sobre las poblaciones de hongos y bacterias actuaría incrementándoles su virulencia. Las poblaciones no evolucionan de forma aislada, sino que lo hacen continuamente unas respecto a otras. Dentro de cada ecosistema, las especies y los nichos están en continua reorganización: a medida que se mueve cada nube de población, influye en las otras nubes, altera su contexto y las empuja suavemente en nuevas direcciones[20]. El espacio genético está lleno de nubes de población que se impulsan a sí mismas, al mismo tiempo que también se empujan entre sí en nuevas direcciones. Este es el principio de la recurrencia operando a nivel de muchas especies en interacción.
§. Una receta para la evolución
En el capítulo anterior vimos que en el corazón de la selección natural subyace un doble bucle de realimentación entre el refuerzo y la competencia. Estos dos bucles interconectados están alimentados por un equilibrio entre la variabilidad y la persistencia de la población, lo que conduce a un incremento de la capacidad para sobrevivir y reproducirse.Pero esta visión de la evolución da por sentado algunas cosas. Supone que tenemos una población de paquetes espaciales integrados llamados individuos. Hemos visto que la individualidad depende de la cooperación, que es tanto un resultado como un ingrediente de la selección natural. Al reunir varios componentes, la cooperación también les permite combinarse de muchas maneras, generando muchas posibilidades. Este principio de riqueza combinatoria es lo que hace que el mundo y sus organismos estén tan preñados de posibilidades.
La interacción entre estos seis principios —variabilidad de la población, persistencia, refuerzo, competencia, cooperación y riqueza combinatoria— proporciona el fundamento de la evolución. Pero el motivo por el que se genera un mundo vivo tan rico y diverso tiene que ver con la recurrencia, nuestro séptimo principio. Cada adaptación desplaza la posición de los objetivos y configura el escenario para que aparezcan más adaptaciones. En lugar de disminuir a medida que las adaptaciones se diseminan en una población, la competencia se incrementa a medida que aparecen nuevas apuestas, instando a la población a satisfacer estándares más elevados. A medida que las poblaciones se dividen, divergen y se reencuentran, el mismo proceso está teniendo lugar entre muchas especies, cada una rivalizando con las otras por un lugar sostenible en un ecosistema complejo. El contexto para la evolución no está fijado, sino que cambia continuamente y se redefine mediante el propio proceso evolutivo. Dado que la evolución cambia continuamente y redefine su propio contexto, nos podríamos preguntar cuál fue el primer contexto evolutivo. La vida se originó hace unos 3800 millones de años, pero no se conocen las circunstancias precisas que condujeron a las primeras formas vivas. Sin embargo, es poco probable que hubiera un comienzo claramente definido porque, como hemos visto, la evolución implica numerosos bucles de realimentación, en cuya naturaleza está no tener un comienzo claro[21]. Los agregados moleculares toscos con una capacidad elemental para replicarse podrían haber sido una forma primitiva de cooperación y persistencia. Estos agregados podrían entonces haberse reforzado mediante la replicación, lo que finalmente da lugar a la competencia por los escasos recursos. La variabilidad que surge durante la reproducción se vería sometida entonces a más refuerzo y competencia, lo que conlleva un mayor nivel de cooperación. A medida que se incrementa la cooperación, los componentes reunidos se podrían combinar de distintas maneras, lo que conduce a una mayor riqueza combinatoria y, por lo tanto, a una mayor diversidad de posibilidades. A medida que se repite este proceso, habrían emergido las primeras formas que podríamos reconocer como seres vivos. Por lo tanto, el contexto de la vida primigenia podría haber surgido por una serie de bucles de realimentación y de interacciones, y no como una etapa nítida.
Cualesquiera que fueran los acontecimientos que condujeron a los primeros atisbos de vida, no se encontrará en ellos la riqueza de la vida, como tampoco se encontrará la riqueza de la pintura en los primeros garabatos humanos en la arena. Más bien, el esplendor de la vida reside en el modo en el que se han transformado profundamente estos atisbos iniciales. A medida que fueron surgiendo las primeras nubes de población, se incitaron a sí mismas a recorrer el espacio genético. Algunas se dividieron para crear nuevas nubes, mientras que otras se extinguieron, pero sus periplos por el inmenso espacio de posibilidades genéticas fueron incesantes, y gracias a ellos surgió la gran gama de patrones y formas vivas.
Nuestros siete principios —variabilidad de la población, persistencia, refuerzo, competencia, cooperación, riqueza combinatoria y recurrencia— y sus interacciones proporcionan la fuerza impulsora para estos periplos, que conducen a la notable variedad de organismos que observamos hoy en día. A esta colección de siete principios y a su forma de trabajar juntos lo he denominado la receta creativa para la vida, que está cimentada en el mundo físico, y descansa en la acción de fuerzas físicas y en las limitaciones de un mundo finito. Por eso la vida es una manifestación importante, pero muy peculiar, que surgió cuando los ingredientes de la receta creativa para la vida se reunieron por primera vez hace varios miles de millones de años.
La evolución es un proceso de transformación que ha hecho posible otros tipos de transformación: que de los huevos se desarrollen adultos, que los recién nacidos se conviertan en adultos sofisticados y que las sociedades tribales se transformen en civilizaciones complejas. ¿Qué tienen que ver estas últimas transformaciones con la evolución? Para responder esta pregunta debemos mirar cómo funcionan estos otros tipos de transformaciones. Comenzaremos con el desarrollo, la primera de las transformaciones que apareció en la evolución.
Capítulo 3
Conversaciones de un embrión
§. Principios de TuringEl cartero llama a su puerta y le tiende un sobre que contiene una invitación en la que pone: «Tenemos el placer de invitarle a una merienda en honor de Alan Turing». Usted aloja en su casa a un amigo que se jacta de su cultura general, por lo que le muestra la invitación. «Ah sí, Turing. Creo que era uno de los tíos que inventó el ordenador. También ayudó a descifrar el código secreto utilizado por los alemanes durante la Segunda Guerra Mundial (creo que se llamaba el código Enigma). Se suicidó cuando era muy joven en los años cincuenta (mordió una manzana untada de cianuro). Me parece una invitación interesante. Yo que tú, iría». Llega a un gran edificio en medio de Manchester. Después de mostrar la invitación en la puerta, le conducen a una gran habitación y le dan una taza de té. Hay mucha gente dando vueltas sola, pero finalmente alguien se le acerca.
§. Formación de patrones en una célula
§. Encendido y apagado de los genes
§. Una lucha molecular
§. El estudio de los gradientes
§. Una forma común
—Hola, Soy Carlos. ¿Sabe de qué va esto?
—Lo siento. No tengo ni idea. Me llegó esta invitación sin esperarlo.
Alguien más le oye y se une.
— ¿A usted también? Sí que es muy extraño. Soy un matemático y trabajo en el mismo departamento que Turing cuando estaba aquí en Manchester, pero no tengo ni idea de lo que pasa hoy.
Unas cuantas personas más se unen a la conversación, pero ahora su grupo se está haciendo muy grande y no alcanza a oír lo que dice cada persona por encima del ruido general de la habitación. Por lo tanto, poco a poco se va situando en el borde y vuelve a quedarse solo. A estas alturas ya se han formado montones de grupos con personas que conversan animadamente. Se siente demasiado visible, por lo que acaba uniéndose a otro grupo de personas y entra en la conversación. La merienda continúa en esta dinámica, con muchos agregados de personas charlando. A pesar de que los individuos de cada grupo cambian con el tiempo, el patrón de agregados permanece. Turing se hubiera sentido feliz.
Durante su vida, Turing se interesó por descubrir las relaciones matemáticas simples que subyacen a los patrones[22]. Sus primeras investigaciones mostraron que nuestra capacidad para solucionar los problemas es un tipo de patrón que se puede expresar matemáticamente. Esta idea le condujo al concepto de máquinas que solucionan problemas, más conocidas como ordenadores. También trabajó en cómo desenmarañar el patrón de símbolos en los mensajes codificados durante la guerra. Hacia el final de su carrera, Turing se fijó en los patrones que surgen durante el desarrollo biológico. Con desarrollo me estoy refiriendo al proceso mediante el cual una única célula, el huevo fecundado, es capaz de transformarse en un organismo pluricelular complejo. Como era típico de Turing, simplificó el problema hasta llegar a la esencia de esta transformación.
Veamos la figura 11, en la que se muestra un patrón de manchas espaciadas que no se consiguió dibujándolas cuidadosamente a mano, sino automáticamente con la aplicación de algunas reglas matemáticas simples. Alan Turing formuló por primera vez estas reglas en 1952 para demostrar que las moléculas son capaces de organizarse por sí solas[23]. El tono de gris del diagrama refleja la concentración de una molécula: el negro para las regiones muy concentradas (donde abunda la molécula), y el blanco para las áreas donde la concentración es más baja. Podemos ver que las moléculas se agrupan en regiones y forman áreas espaciadas de concentración elevada, como los agregados de conversación en la merienda, en la que a pesar de que nadie fue informado de dónde formar los grupos, estos afloraron espontáneamente por la interacción entre las personas. De igual forma, las regiones de concentración molecular elevada de la figura 11 surgen a partir de una distribución inicial uniforme de moléculas y su interacción posterior. Al definir las reglas de interacción, Turing destacó algunos principios clave gracias a los cuales se pueden transformar los patrones desde distribuciones inicialmente simples a organizaciones más complicadas. Como estas reglas se referían a moléculas, Turing pensó que podrían explicar la manera en que una estructura inicial aparentemente simple, el huevo fecundado, podría gradualmente convertirse en un adulto más complejo. Vamos a estudiar de más cerca los componentes principales del sistema de Turing. En el resto del capítulo utilizo algunos términos con los que también se describe la evolución con la idea de resaltar los elementos fundamentales que ambos comparten.

Figura 11. Patrón obtenido al aplicar las reglas de Turing.
§. Principios de Turing
Si colocamos una bolsita de té en una taza de agua caliente, el agua comienza a ponerse marrón a medida que el té se disuelve y se extiende desde la bolsita al resto de la taza[24]. Este proceso de difusión implica una progresión suave del color marrón. No obstante, si pudiéramos acercarnos para observar cada molécula de agua y de té, no observaríamos un movimiento suave, sino frenesí y caos: las moléculas se agitan continuamente por la solución y chocan con otras al azar. Este comportamiento es una consecuencia de la regla física de que las cosas que se encuentran próximas tienden a interaccionar con más fuerza: cuando dos moléculas en solución se acercan demasiado, suelen repelerse o rebotar. Tenemos dos niveles de comportamiento: a escala de la taza de té vemos una progresión suave del color marrón, mientras que a escala molecular vemos frenesí y caos.Estos dos niveles de comportamiento se pueden relacionar con las ideas de la estadística. Tal y como vimos en el capítulo 1, en la estadística intervienen los conceptos de población y variabilidad. En este caso, la población es la colección de moléculas de agua y té de la taza. La variabilidad implica que cada molécula se mueve y choca al azar. Incluso aunque cada molécula siga una trayectoria aleatoria, las de té tienden, de media, a alejarse de su origen, la bolsita, porque no hay nada que las retenga cerca de ella. Así pues, probablemente se alejen después de un rato, incluso si se mueven en direcciones aleatorias entre las colisiones. Por un motivo parecido, un borracho que camine al azar después de salir de casa es muy poco probable que permanezca cerca de ella. El resultado global de los movimientos aleatorios de las numerosas moléculas de té es que se difunden poco a poco por la taza. Dado que el número de moléculas y de colisiones es tan enorme, al tomar la población en conjunto observaremos una progresión suave del color marrón: del caos de los movimientos de cada molécula emerge la difusión regular. Al igual que con otros procesos estadísticos, la difusión no es una propiedad de un suceso único tomado de forma aislada (los movimientos de cada molécula entre las colisiones ocurren en direcciones aleatorias), sino el resultado colectivo de una población.
Las moléculas de té en difusión tienden a alcanzar una distancia promedio en un tiempo determinado. Esto significa que podemos convertir la difusión en un tipo de regla, una medida de la distancia. Pero a diferencia de las reglas que solemos utilizar para medir, esta es estadística y funciona de acuerdo con el comportamiento de la población de moléculas.
Además de la difusión, las colisiones moleculares aleatorias acabarán provocando otro tipo de suceso: una reacción química. Dos moléculas pueden encontrarse y unirse para formar una nueva molécula, igual que dos personas pueden tropezarse y comenzar a conversar. Como las reacciones químicas dependen de las colisiones aleatorias, la velocidad a la que se producen depende de la abundancia, o concentración, de las moléculas implicadas, dependencia que a veces recibe el nombre de ley de acción de masas. Surge como el efecto medio de muchas colisiones al azar dentro de una población de moléculas y, por lo tanto, se trata de otra propiedad estadística. Estas dos formas de regularidad estadística (velocidad de reacción química y de difusión) proporcionan los cimientos del sistema de formación de patrones de Turing, motivo por el cual a veces se le denomina sistema de reacción-difusión.
El que haya demasiada variación puede volverse un problema si queremos generar un patrón: en nuestra merienda no se formarán grupos si cada uno corre de un sitio a otro alocadamente. Por lo tanto, además del movimiento de la población de moléculas, necesitamos un elemento que dé estabilidad. En el caso del sistema de Turing, los productos de las reacciones químicas tienen que ser suficientemente estables para perdurar durante algún tiempo, y las moléculas no deben agitarse ni mezclarse entre sí con demasiada rapidez. Se necesita que haya suficiente variabilidad (movimiento aleatorio de las moléculas) para permitir que empiecen a formarse los patrones, pero no tanta que acabe disipándolo todo. Además de variabilidad en la población, se necesita persistencia.
Una característica clave de una merienda es que una vez que dos personas comienzan a conversar, a las demás les gustará unirse. La conversación fomenta más conversación y, por lo tanto, se estimula a sí misma de forma local. Turing propuso que su sistema molecular tendría un tipo equivalente de refuerzo. Además de participar en las reacciones, algunas moléculas catalizan o favorecen las reacciones. Turing se fijó en lo que ocurriría si el producto de la reacción fuera a su vez una molécula catalizadora. En otras palabras, supongamos que tenemos un catalizador que refuerza una reacción y que el producto de la reacción incrementa la cantidad de catalizador. La molécula catalizadora estimularía su propia producción, esencialmente favoreciéndose a sí misma. Si su concentración fuera ligeramente mayor en un área, tendería a producirse en aún más cantidad en dicha posición. Se trata de un sistema que favorece disparidades y que amplifica las diferencias en vez disiparlas.
Si todo lo que tenemos es refuerzo o autopromoción, el sistema en conjunto simplemente se estimularía a sí mismo para alcanzar concentraciones moleculares cada vez más altas en vez de producir un patrón ordenado. Esto nos lleva a nuestro siguiente principio: la competencia. En el caso de la merienda, la tendencia a que las personas se junten no es constante: a medida que un grupo se hace más grande, se vuelve más difícil escuchar lo que otros dicen por encima del ruido de las demás conversaciones, por lo que las personas tenderán a abandonar los grupos después de que alcancen un determinado tamaño. Cada grupo es víctima de su propio éxito, y por eso terminaremos con muchos grupos de conversación independientes en vez de con un gran conglomerado de personas. En este ejemplo, el aspecto auto limitante del tamaño del grupo surge porque las personas compiten por escuchar lo que se dice. Otra fuente de competencia es la lucha por hablar: a medida que un grupo se hace más grande, encontraremos menos oportunidades para contribuir, con lo que podríamos decidir que lo abandonamos.
Turing también invocó una forma de competencia a la hora de generar estructuras moleculares. La competencia surgirá, entre otros motivos, por la limitación natural de la reacción química: la producción de una molécula en una reacción implica el consumo de otras moléculas, denominadas sustratos, que están presentes en la solución. Cada vez que se genera un producto de forma auto inducida, se consumen moléculas de sustrato. Dado que el suministro de sustratos no es infinito, el proceso continuará hasta que comiencen a agotarse. La molécula auto inducida se convierte en una víctima de su propio éxito: cuanta más se fabrica, menos sustrato queda disponible para su síntesis y empiezan a competir entre sí por el poco sustrato que va quedando. La competencia se va intensificando a medida que se consume más sustrato, lo que retrasa todavía más el incremento de la molécula auto inducida.
El proceso se puede resumir mediante dos bucles de realimentación interconectados (figura 12). La molécula auto inducida forma un bucle de realimentación positiva porque estimula su propia síntesis (refuerzo). El bucle de realimentación negativa indica el efecto estrangulador por la limitación del sustrato sobre el sistema (competencia). Estos bucles operan en el contexto de la difusión: tanto las moléculas auto inducidas como las de sustrato están en continua agitación por la solución, lo que les permite diseminarse y participar en reacciones químicas. Turing demostró que, según lo rápido que difundan las moléculas auto inducidas y de sustrato, a veces acabarán formándose regiones espaciadas de concentración elevada, como en la figura 11. Esto ocurrirá incluso si comenzamos con una distribución uniforme de moléculas. Al igual que con las conversaciones en la merienda, la autoinducción puede conducir a un refuerzo local, mientras que la competencia disgrega las cosas y ayuda a esparcirlas. Turing había demostrado que con algunas reglas simples se podría conducir a la transformación de una distribución inicialmente uniforme en un patrón de manchas más complejo.
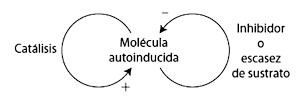
Figura 12. Interacción entre el refuerzo (bucle positivo) y la competencia (bucle negativo) en un sistema de Turing.
En este caso, la limitación del ascenso de la molécula auto inducida viene provocado por la síntesis de un inhibidor y no por la competencia por una cantidad escasa de sustrato. No obstante, el principio fundamental de un bucle de realimentación negativa impulsado por una entidad auto inducida es muy similar. Utilizo competencia como término general para describir ambas formas de bucle de realimentación negativa, como limitaciones ocasionadas por la inhibición o como escasez de un componente.
En la raíz de estos sistemas de formación de patrones encontramos algunas similitudes formales con lo que describimos para la selección natural en el capítulo 1. En ambos casos encontramos un doble bucle de realimentación con refuerzo y competencia, alimentado por un equilibrio entre la persistencia y la variabilidad de la población. A diferencia de la selección natural, en el sistema de Turing no intervienen poblaciones de organismos que se reproducen durante generaciones, sino que afecta a poblaciones de moléculas que interaccionan durante mucho menos tiempo. En él intervienen un conjunto de principios parecidos a los de la selección natural, pero bajo una nueva apariencia. El resultado neto no es la evolución de los organismos, sino la transformación de los patrones dentro de cada organismo.
En el momento en el que Turing propuso sus ideas, se sabía poco sobre los mecanismos moleculares que subyacen a la formación de patrones. El modo en el que trabajan los genes tampoco se conocía bien, y ni tan siquiera se había descubierto la estructura del ADN. Su esquema fue más bien especulativo, con muy pocas pruebas directas a favor o en contra. Sin embargo, en las pasadas décadas hemos aprendido mucho más sobre cómo transcurre realmente el desarrollo[26]. Aunque estos hallazgos más recientes se describan a veces con términos diferentes a los que Turing utilizó, veremos que implican la misma clase de principios a los que apeló.
§. Formación de patrones en una célula
Para ilustrar la imagen que ha aflorado de los descubrimientos más recientes, comencemos con un caso de formación de patrones en el mundo unicelular. Cada vez que se reproduce una bacteria de la especie Escherichia coli, se forma un patrón: se genera un tabique en el centro de la célula que la divide en dos. ¿Cómo se las apaña E. coli para trazar una línea por el centro? La posición del tabique depende de una interacción entre dos proteínas, denominadas MinD y MinE[27]. Estas proteínas custodian los dos extremos de la célula, e impiden que se forme un tabique en ellos, con lo que garantizan que solo se formará en el centro. Si contempláramos a MinD y a MinE durante un tiempo, veríamos que están muy concentradas en un extremo de la célula y a continuación, unos minutos después, la concentración se eleva en el otro extremo (figura 13). Se comportan como policías con un ritmo regular, moviéndose hacia atrás y hacia delante para garantizar que no se forme ningún tabique cerca de los extremos de la célula. ¿Qué guía este curioso comportamiento de las proteínas Min?Comencemos con MinD. Si observamos de cerca la ubicación de la proteína MinD, veremos que, además de difundirse con libertad dentro de la célula, también es capaz de unirse a la membrana celular, con lo que permanecerá ahí durante cierto tiempo. Esta unión a la membrana también favorece que otras moléculas MinD se unan cerca de ella. En otras palabras, MinD refuerza la fijación de sus semejantes. Podríamos esperar que, al final, todas las moléculas de MinD estén fijadas a la membrana. Sin embargo, esto no ocurre debido al efecto competidor de MinE, la otra proteína del equipo: MinE se une a las moléculas de MinD de la membrana y hace que se desprendan y vuelvan al interior de la célula. Esto significa que MinD se convierte en una víctima de su propio éxito: cuanto más se une a la membrana, más MinE acude a esa posición y más MinD se desprende entonces. Podemos resumir esta relación con nuestro conocido diagrama del doble bucle de realimentación (figura 14). El bucle positivo representa la fijación de MinD a la membrana y el fomento de la fijación de más moléculas de MinD. El bucle negativo representa la atracción que MinD ejerce sobre MinE hacia la membrana, lo que la hace desprenderse. Tenemos todos los ingredientes necesarios para que aparezca un patrón. Las simulaciones por ordenador demuestran que, con las afinidades moleculares y las tasas de difusión adecuadas, este sistema es capaz de generar el patrón oscilante observado para las proteínas Min, en el que la concentración elevada avanza y retrocede continuamente desde un extremo de la célula hasta el otro cada cinco minutos.

Figura 13. Las proteínas Min custodian los extremos de una célula de E. coli.
Figura 14. Interacción entre el refuerzo (bucle positivo) y la competencia (bucle negativo) en la formación del tabique en la célula de E. coli.
Figura 15. Ejemplos de tipos celulares de los animales (arriba) y de las plantas (abajo).
§. Encendido y apagado de los genes
Recordemos que un gen es un tramo de ADN. En muchos casos podemos dividirlo grosso modo en dos partes: una región reguladora y una región codificante (figura 16). Fijémonos primero en la región codificante, pues contiene la información necesaria para especificar la secuencia de aminoácidos de una proteína. La información no se lee directamente del ADN, sino que primero se copia, o se transcribe, para producir otro tipo de molécula llamada ARN, cuya estructura es similar al ADN, pero solo tiene una hebra en lugar de dos. Si pensamos que el ADN del núcleo de una célula es un conjunto de libros, en el que cada gen es una página, entonces el ARN corresponde a una fotocopia de parte de una página (la región codificante). Un mismo gen puede sintetizar muchas moléculas de ARN, al igual que se hacen muchas fotocopias de la misma página. Tras la copia, las moléculas del ARN salen del núcleo hacia el líquido celular que lo rodea, o citoplasma, donde su información se traduce en la secuencia de aminoácidos de una proteína. A continuación, la proteína se pliega de una forma particular, según su secuencia. El proceso completo se ha resumido con la expresión «El ADN hace el ARN que hace la proteína».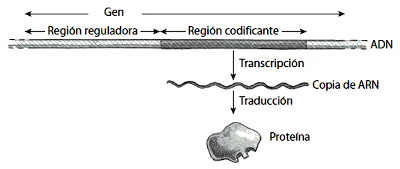
Figura 16. Gen típico que muestra cómo se transcribe la información contenida en el ADN para sintetizar el ARN, que a su vez se traduce para sintetizar una proteína.
Esta pregunta nos lleva a la región reguladora. La figura 17 nos muestra un diagrama simplificado de la región reguladora de un gen, donde podemos ver que tiene varias proteínas unidas a ella, simbolizadas en forma de manzanas y limones, que se denominan proteínas reguladoras, o proteínas maestras. Las proteínas reguladoras reconocen pequeños tramos del ADN y se fijan a ellos. La proteína reguladora con forma de manzana se fija a la secuencia de ADN simbolizada por tramos gris oscuro, mientras que la proteína reguladora con forma de limón se fija a las secuencias gris claro. Una vez que se fijan al ADN, las proteínas reguladoras activan o reprimen la transcripción del gen, con lo que hacen subir o bajar la actividad del gen. Lo consiguen mediante la alteración de la maquinaria de copia del ARN que se acopla al gen y lo transcribe. Imaginemos que la proteína reguladora con forma de manzana favorece la transcripción del gen cuando se fija, por lo que lo enciende. Entonces, la proteína manzana sería un activador del gen. Por el contrario, la proteína reguladora con forma de limón sería un inhibidor, que reprime al gen cuando se fija a él y tiende a apagarlo. El nivel de encendido o apagado del gen dependerá del equilibrio entre las proteínas activadora e inhibidora que se unan a la región reguladora.
Podríamos preguntarnos de dónde vienen las proteínas reguladoras (los limones y las manzanas): resulta que están a su vez codificadas por otros genes (la proteína limón por un gen y la proteína manzana por otro diferente). Por lo tanto, hay genes que codifican proteínas que, a su vez, regulan a otros genes. Como veremos, una consecuencia de estas interacciones entre los diferentes genes y proteínas es la generación de los patrones.
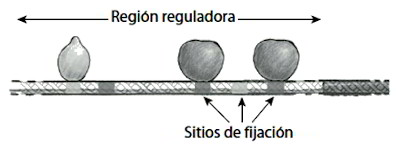
Figura 17. Proteínas fijadas a la región reguladora de un gen.
§. Una lucha molecular
Muchos de los casos mejor conocidos de formación de patrones se encuentran en la mosca del vinagre Drosophila melanogaster. Esta especie resulta cómoda para estudiar mutaciones y genes porque se reproduce rápidamente y se mantiene con facilidad en poblaciones grandes en los laboratorios. Fijémonos en la formación de un tipo determinado de célula, denominada neuroblasto, en una mosca del vinagre en desarrollo[28]. Los neuroblastos son las células que finalmente dan lugar al sistema nervioso de la mosca. El panel superior de la figura 18 muestra una región pequeña del embrión de la mosca en desarrollo, donde se están formando dos neuroblastos. Podemos ver que al principio hay dos grupos de células de color gris claro, y que de cada uno emerge una única célula de neuroblasto (gris oscuro). Esto nos acarrea un problema de formación de patrones básico: ¿cómo se singulariza una célula respecto a sus vecinas para convertirse en un neuroblasto?
Figura 18. Formación de patrones neurales en las moscas del vinagre.
Para ver cómo funciona, fijémonos en la parte izquierda de la figura 19, en la que dos células gris claro interaccionan entre sí. Podemos ver algunas flechas con signos positivos y negativos. El efecto global de estas interacciones equivale a nuestros conocidos bucles de realimentación negativa y positiva. En cada célula, el gen Delta (localizado en el núcleo) se enciende y produce la proteína Delta (por convención, los nombres de las proteínas no se escriben en cursiva, para diferenciarlas de los genes que las codifican). Las moléculas de la proteína Delta, simbolizadas en forma de piruleta, se dirigen a la membrana celular, en la que se insertan con la cabeza hacia el exterior de la célula. Como se están zarandeando por la membrana, pueden acabar colisionando con otra proteína, un receptor, localizada en la célula vecina. El receptor tiene una forma que encaja con la de la proteína Delta. Si Delta se fija al receptor, se enciende una cadena de acontecimientos en la célula portadora de dicho receptor que hace que ciertas moléculas se difundan al núcleo y apaguen el gen Delta (el signo negativo en la figura).
Esencialmente, la síntesis de Delta en una célula supone un gran impacto para su vecina, porque intenta apagar su gen Delta. Si la cantidad de la proteína está razonablemente equilibrada, ambas células apagan respectivamente su gen Delta al mismo nivel. Pero si una célula tiene ligeramente más cantidad de Delta, entonces su concentración se incrementará más (se refuerza) a medida que apaga con más eficacia la de la célula vecina competidora. Finalmente, ganará una célula que entonces contendrá gran cantidad de Delta mientras que las otras expresarán muy poca (figura 19, derecha). He ilustrado la interacción entre dos células, pero si las mismas reglas se extendieran a un grupo pequeño, se puede obtener un resultado parecido y al final una de las células contendrá muchísima más proteína Delta que las otras. Los principios que intervienen son similares a los que aducía Turing, pero en vez de tratar con las reacciones químicas simples en una solución, manejamos cadenas de acontecimientos en las que las células se envían señales unas a otras y los genes se encienden o se apagan.
Figura 19. Señalización intercelular mediante la proteína Delta y su receptor.
§. El estudio de los gradientes
Hemos visto que la interconexión entre bucles de realimentación conduce a la transformación de los patrones, pero quiero ahora ocuparme de otro tipo de mecanismo de formación de patrones que, a primera vista, parece basarse en principios diferentes. La parte superior de la figura 20 muestra una línea de células en la que hay un gradiente de concentración de una molécula, cuya concentración es máxima en el extremo izquierdo (negro) y disminuye poco a poco hasta la concentración más baja de la derecha (blanco). Podríamos imaginar que este gradiente surge porque las moléculas se sintetizan en las células del extremo izquierdo y se difunden hacia el otro extremo. Supongamos ahora que estas moléculas modifican la actividad de un gen, de modo que lo encienden si la concentración de la molécula está por encima de un umbral. Terminaríamos con dos regiones diferentes: una región izquierda en la que el gen está encendido (la concentración de la molécula está por encima del umbral), y una región derecha con el gen apagado (figura 20, panel inferior). La actividad del gen no la veremos como un gradiente, sino como un patrón escalonado: elevada en la mitad izquierda y baja en la mitad derecha. Hemos pasado de un patrón inicial relativamente suave a otro más abrupto. Parece, no obstante, que aquí no necesitaremos los principios de refuerzo ni de competencia puesto que para generar un patrón escalonado nos basta con crear el gradiente por difusión y una concentración umbral que afecte a la actividad del gen. Pero ¿cómo se miden realmente los umbrales? Nos resulta fácil dibujar una línea y decir que lo que esté por encima de ella se comporta de modo distinto de lo que esté por debajo. No obstante, ¿cómo consiguen las células hacer lo mismo? Para responder a esta pregunta, quiero examinar uno de los ejemplos de gradiente mejor estudiados.En la historia que viene a continuación interviene la interacción entre dos genes, denominados Bicoid y Hunchback. En aras de la simplicidad, las proteínas sintetizadas por estos genes las representamos por dos frutas: manzanas para Bicoid y peras para Hunchback. Empecemos por las manzanas. Una hembra de la mosca del vinagre produce a lo largo de su vida muchísimos óvulos, cada uno de una longitud aproximada de un milímetro. Después de la fecundación, el núcleo de los huevos se somete a varias rondas de división que le harán contener muchos núcleos. Es poco habitual que los núcleos se dividan así dentro de una célula, pero solo es de forma temporal; más tarde, cada núcleo se empaquetará en una célula diferente. A medida que se multiplican los núcleos, en el embrión se establece una distribución gradual de la proteína manzana (Bicoid), cuya concentración es elevada en el extremo de la cabeza y baja en el extremo de la cola (figura 21, por ahora ignoraremos la cuestión de cómo aparece este gradiente). En algunos núcleos, la proteína manzana enciende nuestro segundo gen, Hunchback, lo que conduce a la producción de peras. En otras palabras, las manzanas se unen a la región reguladora del gen Hunchback y lo encienden. Esto ocurre principalmente en los núcleos del extremo de la cabeza, donde las manzanas abundan más. El resultado es que las peras también abundan en este extremo (figura 22). Pero advertiremos que al encender la producción de peras, el gradiente de manzanas ha creado un patrón más escalonado de peras: la curva de la concentración de peras es casi plana en el extremo de la cabeza y luego desciende con más brusquedad que el gradiente de manzanas (figura 22, panel inferior). Se trata, pues, de un ejemplo de una distribución gradual de una proteína que da lugar a una gradación más abrupta de otra proteína durante el desarrollo.

Figura 20. Gradiente y umbral que conducen a una actividad génica con un patrón escalonado.
Figura 21. Gradiente de concentración de la proteína Bicoid (manzanas) desde la cabeza a la cola en el embrión de la mosca del vinagre.
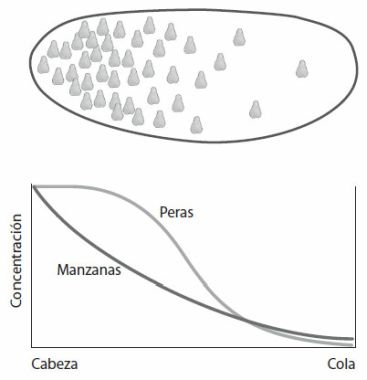
Figura 22. Patrón escalonado de la concentración de la proteína Hunchback (peras) desde la cabeza a la cola del embrión de la mosca del vinagre.
Figura 23. Las proteínas Bicoid (manzanas) se fijan a la región reguladora del gen Hunchback, lo que incrementa su actividad y la concentración de la proteína Hunchback (pera).
§. Una forma común
Todos los ejemplos de formación de patrones que nos hemos encontrado en este capítulo tienen algunas características básicas en común. Cada uno se basa en la interconexión de bucles de refuerzo y de competencia, alimentados por un equilibrio entre la variabilidad de la población y la persistencia: la formación de grupos de conversación en la merienda, el sistema de difusión-reacción de Turing, la división de una célula de E. coli, la lucha molecular por la supremacía de Delta en los neuroblastos y la construcción del gradiente de Bicoid en las primeras fases del embrión de la mosca. Todos ellos se fundamentan en la misma receta básica cuyo resultado global es la refinación de los patrones: mayor agrupamiento de la actividad o delimitación nítida de la distribución. Si nos fijáramos en otros ejemplos de formación de patrones en desarrollo, encontraríamos continuamente los mismos ingredientes básicos. Los genes y moléculas concretos variarán en cada caso, pero los principios serán los mismos. ¿Por qué encontramos siempre la misma organización?Para refinar un patrón molecular es necesario coordinar el incremento de algunos componentes y la disminución de otros. El refuerzo por sí solo no daría lugar más que a una auto estimulación de los componentes hacia concentraciones más y más altas. Por el contrario, la competencia por sí sola provocaría el consumo general de los componentes (sustratos), o la inhibición que los disminuiría en cualquier parte. No obstante, la situación es mucho más interesante cuando van unidos el refuerzo y la competencia: en vez de una subida o bajada generales, algunos componentes suben a expensas de otros. Si nosotros también incorporáramos los efectos de la distancia a través de la señalización o de la difusión moleculares, los componentes se organizarían en el espacio y ocuparían regiones diferentes. Los componentes se harían más abundantes en algunos lugares, pero no en otros. De este modo se puede refinar un patrón, con la delimitación nítida de una frontera o con la activación de un gen en un subconjunto de células.
La interacción entre la variabilidad de la población, la persistencia, el refuerzo y la competencia se puede comparar con la que encontramos con la selección natural en el capítulo 1. En el caso del desarrollo, la variabilidad de la población implica interacciones estadísticas entre numerosas moléculas o células de un individuo. En el caso de la evolución, la variabilidad de la población implica que estén surgiendo mutaciones continuamente, y que se distribuyan por una población de individuos. En ambos procesos se necesita la variabilidad de la población para que se produzca el cambio.
Sin embargo, el exceso de variabilidad es un problema. El grado de variación tiene que permanecer confinado, o si no, los cambios se disiparían en cuanto hubieran cobrado forma. La persistencia se establece durante el desarrollo mediante la estabilidad química y los límites a las mezclas moleculares. Las membranas celulares, por ejemplo, impiden que el contenido de una célula se mezcle con demasiada rapidez con el de sus vecinas. También ayudan a que el contenido de las células, que incluye las proteínas reguladoras, no se disperse y pase a las células hijas, lo que permite que se mantengan los patrones de la actividad génica. De igual forma, la velocidad de difusión intracelular e intercelular de las moléculas señalizadoras está confinada entre unos límites estrictos. En la evolución, la persistencia implica la transmisión fiel de la información de una generación a otra mediante la reproducción y la replicación génica. En el desarrollo y en la evolución, se necesita que la persistencia compense la variabilidad: sin variabilidad no cambiaría nada y sin persistencia no perdurarían los cambios.
La variabilidad de la población y la persistencia son ingredientes esenciales, pero cada una por su cuenta no generan resultados interesantes. Para que aparezca la organización, unos componentes tienen que aumentar a expensas de otros, lo que ocurre por la interacción entre el refuerzo y la competencia. En el desarrollo, el refuerzo se manifiesta cuando las moléculas estimulan su propia síntesis o potencian su actividad. El incremento de estas moléculas se contrarresta con la activación de inhibidores o con la competencia por factores limitantes. Esta relación entre el refuerzo y la competencia proporciona la fuerza impulsora para generar patrones. En la evolución, el refuerzo se basa en que los genes favorecen su propia replicación gracias a que influyen en la supervivencia y en la reproducción. Luego el éxito reproductor incrementa la competencia debido a la escasez de recursos. En ambos casos, lo que conduce a un cambio organizado es el acoplamiento entre el refuerzo y la competencia.
A pesar de las muchas diferencias entre el desarrollo y la evolución, vemos los mismos principios e interacciones. No debería sorprendernos tanto que los principios que utilizo para describir la evolución también se encuentren en mi explicación del desarrollo. Como mencioné al comienzo del capítulo 1, llegué a esta colección concreta de principios porque busqué las características que eran comunes a las diferentes transformaciones de los seres vivos, no solo a una. Lo que quizá sorprenda más es que los puntos de encuentro que hayamos no son superficiales, sino que se encuentran en el centro de cada proceso. Definen las interacciones principales que conducen a las transformaciones en cada caso, lo que sugiere que la evolución y el desarrollo no operan de forma completamente diferente, sino que se trata de manifestaciones distintas de un conjunto común de principios subyacentes a través de los cuales emerge la organización.
En cualquier caso, debemos tener cuidado de no llevar tales comparaciones demasiado lejos. Por ejemplo, mientras que las mutaciones desempeñan una función crucial en la evolución, no existe un equivalente exacto en el desarrollo[11]. Una mutación implica un cambio singular heredable en una gran molécula de ADN; no se parece en nada a las colisiones entre las moléculas de las reacciones y de la difusión. De igual forma, el refuerzo en la evolución implica la replicación, mientras que las moléculas pueden reforzar su acción durante el desarrollo sin copiarse directamente. Las comparaciones pueden dar lugar a confusión en vez de resultar útiles si se toman muy al pie de la letra, como en el ejemplo de la introducción sobre un profesor que comparaba un caballo con el caballo del ajedrez. La guerra y el ajedrez muestran similitudes en la forma y no una correspondencia pormenorizada. De igual forma, no debemos intentar emparejar los componentes de la evolución y del desarrollo con demasiada precisión, pues estamos tratando con una similitud en la forma, no en el detalle, con principios y bucles de realimentación similares. El desarrollo y la evolución utilizan la misma receta básica, pero se visten con dos trajes muy diferentes.
Sin embargo, al dar a conocer los principios básicos del desarrollo, he pasado por alto muchas cuestiones. Cada una de las transformaciones que he descrito supone un contexto previo: una región de células que forman neuroblastos, o un gradiente de la proteína Bicoid en el huevo. Pero ¿cómo se generan estos contextos a sí mismos? ¿Cómo se relacionan unas pocas transformaciones de patrones con el desarrollo de organismos pluricelulares complejos, como los manzanos y los humanos? Para responder a estas preguntas debemos investigar la manera en que tiene lugar el desarrollo.
Capítulo 4
Completemos el cuadro
§. Un aperitivo embrionarioAlgunos de los cuadros de Cézanne parecen inacabados. En El Jardín de Les Lauves (figura 24, lámina 4), Cézanne dejó de pintar después de bosquejar algunas áreas con color. No sabemos por qué dejó el cuadro de este modo: quizá llegó un momento en el que ya no pudo seguir, o quizá le parecía perfecto tal y como estaba. Cualquiera que fuera la razón, el cuadro revela cómo elaboró la textura y las formas. Parece que comenzó con un boceto de color que fue elaborando con la aplicación de otras manchas de color. Cada pincelada de color dependía de los colores que ya estaban en el lienzo y, a su vez, influían en los siguientes colores que aplicaba. La descripción que ofrece Cézanne sobre el cuadro del retrato también da a entender este proceso enormemente interactivo: «Si tejo en torno a tu expresión la red infinita de pequeños azules y marrones que están ahí, que casan entre sí, haré que mires desde el lienzo igual que en la vida real… Una pincelada después de otra, una después de otra».
§. Un esfuerzo cooperativo
§. Riqueza reguladora
§. Se construye sobre el pasado
§. El lienzo en expansión
§. Deformación
§. El lienzo tridimensional
En el capítulo anterior vimos cómo se transforman los patrones durante el desarrollo, cómo a partir de una distribución inicial se producían nuevos patrones de actividad génica, por ejemplo una disposición en manchas o escalonada. Se parece mucho a la manera en que un artista añadiría unas pocas pinceladas más de color a un lienzo para transformar su aspecto. Pero unas pocas pinceladas por sí mismas no son suficientes para crear un patrón complejo. El cuadro tiene que construirse colocando color tras color, pincelada tras pincelada. El desarrollo de un embrión no implica tan solo una o dos transformaciones, sino que en su lugar interviene la acumulación de muchos cambios, cada uno construido sobre el anterior. ¿Cómo funciona este proceso de elaboración continua?

Figura 24. El jardín de Les Lauves, Paul Cézanne, 1906. Véase la lámina 4.
§. Un aperitivo embrionario
Pensemos en un embrión en desarrollo como si fuera un lienzo en el que se habrán de colocar los colores y los patrones. Como vimos en el capítulo anterior, en las primeras etapas del embrión de la mosca del vinagre hay un gradiente de proteínas reguladoras desde la cabeza a la cola, simbolizado por manzanas, lo que equivale a tener una aguada de color inicial en nuestro lienzo, pongamos verde manzana, que gradualmente se va desvaneciendo de un extremo al otro (figura 25, arriba). Este gradiente de manzanas se aprovecha luego para generar un patrón escalonado de peras, que se produce mediante la fijación de las proteínas manzana a la región reguladora del gen de la pera, con lo que queda encendido a distinta intensidad a lo largo del embrión (figura 25, centro). Ahora tenemos dos colores en nuestro lienzo: un gradiente suave de verde manzana y un patrón más escalonado de verde pera. Como en el cuadro, el segundo color se ha colocado influido por el primero.
Figura 25. Refinamiento y elaboración de los patrones en las primeras etapas de un embrión de la mosca del vinagre.
Lo que sigue es una interacción complicada entre los diversos genes de las frutas, parecida a la de los colores en un lienzo. Al igual que cada pincelada aplicada por un artista depende de otras pinceladas e influye en las siguientes, cada gen contribuye y responde a otros genes, porque no solo codifican una fruta (proteína reguladora), sino que también tienen una región reguladora a la que se fijan las frutas. El resultado es que algunos genes y sus proteínas reguladoras tienden a reforzarse mutuamente la actividad, mientras que otros compiten. Los nuevos patrones comienzan a emerger por una batalla territorial: las bananas están confinadas en un extremo, y después van bandas de limones, mangos, etc., hasta las piñas en el otro extremo (figura 25). Los principios que gobiernan este proceso no son diferentes de los que encontramos en el capítulo anterior: los patrones se transforman poco a poco a través del zarandeo molecular, la persistencia, el refuerzo y la competencia. En este punto, sin embargo, se están produciendo muchas interacciones en paralelo cuyo resultado global es que después de haber comenzado con un mero gradiente de verde manzana, el patrón de nuestro lienzo se ha ido elaborando hasta producir diferentes bandas de color. Este proceso no está gobernado por ningún artista externo, sino que ha emergido gracias a las interacciones moleculares.
§. Un esfuerzo cooperativo
Por debajo de estos acontecimientos en el embrión está el modo en el que las distintas proteínas reguladoras (idealizadas aquí por frutas) interaccionan cuando se fijan a las regiones reguladoras de los genes. Cada región reguladora puede contener diez o más sitios diferentes a los que se pueden fijar distintas proteínas reguladoras. En un gen, por ejemplo, podría tenerlos para manzanas, peras, bananas, limones y mangos, y actuaría como un imán estadístico que tenderá a juntar estas frutas. Al encontrarse muy cerca unas de otras, resulta fácil que se empujen e interaccionen unas con otras, mediante lo cual se determinará si el gen se enciende o se apaga. Al juntar frutas, las regiones reguladoras alientan las interacciones, con lo que no podremos predecir lo que hará una proteína reguladora sin tener en cuenta cómo actúa respecto a otras. Para que el desarrollo continúe de forma ordenada, el resultado global de estas interacciones es la activación o inhibición coordinada de los genes a medida que se desarrolla el embrión. Este es el principio de cooperación que actúa sobre las regiones reguladoras del gen.La cooperación no solo actúa sobre las regiones reguladoras. Recordemos que a medida que se desarrolla el embrión de la mosca del vinagre, se dividen los núcleos en su interior. El huevo fecundado comienza con un único núcleo, pero cuando se han formado las bandas de bananas, de limones y de otras frutas, el embrión ya contiene muchos núcleos, cada uno con su propia copia del ADN. La interacción entre los núcleos se produce al estar unos muy cerca de otros. Las proteínas reguladoras sintetizadas por un núcleo, pongamos limones, se trasladan a los núcleos más próximos e influyen en las proteínas reguladoras que se sintetizarán. Esta conversación entre los núcleos cercanos es otra forma de cooperación que permite que el desarrollo prosiga de una manera organizada.
Tener varios núcleos, en vez de uno solo, dentro de una célula es una peculiaridad de las primeras etapas del embrión de la mosca del vinagre. En la mayoría de los animales y plantas, las células de las primeras etapas del embrión se dividen, y sus núcleos lo hacen al mismo tiempo para producir varias células, cada una con una membrana externa y un único núcleo. Estos embriones pluricelulares presentan patrones con la misma clase de principios que la mosca, salvo que se necesitan otras moléculas para enviar y recibir señales a través de las membranas (con un funcionamiento parecido a la señalización Delta y el sistema receptor descrito en el capítulo anterior. De nuevo encontramos la cooperación local, pero esta vez son las células cercanas en el embrión las que interaccionan entre sí. Las interacciones de este tipo entre las células también tienen lugar en las etapas más tardías de la mosca del vinagre, cuando los numerosos núcleos de su embrión acaban rodeados por membranas y forman células independientes. El desarrollo es siempre un esfuerzo cooperativo de interacciones entre las células cercanas, los núcleos y las moléculas reguladoras.
§. Riqueza reguladora
Se obtienen diferentes resultados cuando interaccionan entre sí varios componentes diferentes. El inmenso abanico de moléculas del universo se forma por distintas combinaciones de tan solo una centena de átomos, y todos los genomas en la Tierra surgen por combinaciones diferentes de tan solo cuatro bases: A, G, C y T. Este principio de riqueza combinatoria también se aplica a la regulación génica. Hasta ahora hemos tratado con nueve proteínas reguladoras, simbolizadas por manzanas, peras, bananas, etc. Pero no es más que la punta del iceberg: cada planta o genoma animal puede codificar cientos, o incluso miles, de proteínas reguladoras. El genoma humano, por ejemplo, contiene unos 25.000 genes, de los que se piensa que unos 2.500 codifican proteínas reguladoras que pueden encender o apagar a otros genes. Es decir, el genoma humano codifica cerca de 2.500 tipos diferentes de frutas.Para ver a lo que conduce esto, supongamos que un genoma codifica 1000 tipos diferentes de proteínas reguladoras. En un momento dado, una célula determinada, o un núcleo con su citoplasma circundante, sintetiza tan solo una fracción de estas proteínas, digamos el 10%. Esto significa que cada célula produce 100 de las 1000 proteínas reguladoras posibles, una selección de 100 frutas de las 1000 posibles. Otra célula podría tener una colección diferente, una combinación distinta de 100 frutas. ¿Cuántos tipos diferentes de células, en términos de producción de proteínas reguladoras, podríamos llegar a tener con este sistema? La respuesta es alrededor de 10 300 (un uno seguido por 300 ceros), que es un número colosal. Estamos tratando con un enorme hiperespacio de 100 dimensiones (la selección de frutas), cada uno con 1.000 posibilidades. Este es el principio de la riqueza combinatoria aplicada a las proteínas reguladoras.
Pero la riqueza combinatoria no se detiene aquí: si tenemos diferentes tipos de células en cercana proximidad, se abren aún más posibilidades. Podemos disponer las células de diversas maneras, tal como podemos ordenar las piezas del ajedrez también de diferentes maneras sobre el tablero. Sin embargo, a diferencia del ajedrez, que tiene solo doce tipos de piezas (seis negras y seis blancas), tenemos 10300 posibilidades con las que jugar, y en vez del tablero con 64 escaques, podemos tener muchos miles o millones de células, un número de combinaciones posibles que no somos capaces ni de imaginar. Tenemos un enorme hiper hiperespacio de posibilidades de tipos de células y de organizaciones. A este gran espacio de posibilidades lo denomino espacio del desarrollo.
Aunque el espacio del desarrollo es inimaginablemente grande, podemos visualizarlo como un espacio tridimensional muy grande. Al igual que hemos representado la evolución como poblaciones que viajan a través de un enorme espacio genético (capítulo 2), podemos pensar en el desarrollo como un embrión que viaja por un enorme espacio del desarrollo. Tras la fecundación, el embrión está formado por una única célula con una combinación concreta de proteínas reguladoras que ocupan una posición en el espacio del desarrollo. A continuación se activan determinadas proteínas reguladoras en distintas regiones del embrión a medida que se dividen el núcleo y las células, lo que corresponde a un embrión que sigue una ruta concreta por el espacio del desarrollo. El ejemplo del embrión de la mosca adentrándose en su serie de patrones afrutados representa el comienzo de este periplo. El embrión de la mosca está negociando un estrecho sendero a través del enorme espacio de posibilidades reguladoras, igual que Cézanne pintando un cuadro representa un peregrinaje por el enorme espacio de posibilidades del color. Hasta el momento hemos contemplado unas pocas etapas de este recorrido, o unos pocos ejemplos de la manera en que un patrón de actividades génicas se transforma en un patrón ligeramente diferente. Para ser más conscientes de lo que conlleva este recorrido, avancemos un poco más en el episodio de la mosca del vinagre.
§. Se construye sobre el pasado
En la parte superior de la figura 26 podemos observar el embrión de la mosca del vinagre explicado antes, con las proteínas banana en el extremo de la cabeza y las piñas en el extremo de la cola. Durante las siguientes etapas del desarrollo ocurren varias cosas: una es que los núcleos del embrión se rodean de membrana celular y se forman células individuales. Otra es que comienza a aparecer un patrón repetitivo de actividad génica, como se muestra a la derecha de la figura 26. Se sintetizan varias proteínas reguladoras nuevas, que se simbolizan con tres frutas: fresas, mandarinas y uvas. Donde antes solo había un color, que representa las bananas, por ejemplo, ahora hay una tira delgada de fresas, seguida de una tira de mandarinas, una tira de uvas, luego otra tira de fresas, mandarinas y uvas. El mismo tema fresa-mandarina-uva se repite una y otra vez a lo largo del embrión. Estas proteínas las sintetizan otro conjunto de genes (conocidos como genes de la polaridad de los segmentos). Su patrón repetitivo de actividad se genera al aprovechar la distribución inicial de bananas, limones, etc., según los principios que ya hemos encontrado antes. Pero en este caso, los genes se encienden en posiciones repetidas a lo largo del embrión.Al mismo tiempo que se establece este tema repetitivo, se produce otro conjunto de proteínas reguladoras, como se muestra a la izquierda de la figura 26. Este patrón de proteínas no es tan diferente de nuestro patrón anterior de bananas, limones, etc. La principal diferencia reside en que los límites entre las regiones son más nítidos y están mejor definidos. Estas proteínas reguladoras las sintetiza un nuevo conjunto de genes (denominados genes Hox).
Ahora imaginemos que se superponen los dos patrones, como se muestra en el embrión de la parte inferior de la figura 26. El resultado es un tema con variaciones: un patrón repetitivo combinado con modulaciones provocadas por la progresión de dominios definidos con nitidez desde la cabeza a la cola. Este patrón de superposiciones proporciona un armazón básico para la mosca. Cada mosca adulta tiene catorce segmentos, de los que algunos forman la cabeza, otros la parte central (tórax) y el resto la sección de la cola (abdomen). Los segmentos varían de uno a otro: un segmento lleva antenas, otro lleva patas, otro lleva alas, y así sucesivamente. Tenemos un tema repetitivo, el segmento, y variaciones debidas a que los segmentos son distintos. Esta organización se remonta al patrón de proteínas reguladoras de las primeras etapas del embrión, nuestro patrón de solapamiento de frutas. Dicha combinación inicial de proteínas reguladoras define la distribución modular de la mosca y establece los patrones de actividad génica que conducen a la organización segmentada del adulto. Las mutaciones que inactivan los genes que codifican las proteínas reguladoras dan lugar a moscas que se desarrollan sin algunos segmentos, o con un tipo de segmento reemplazado por otro[33].
Figura 26. Interacciones entre los temas y las variaciones en un embrión de la mosca del vinagre.
¿Qué impulsa al embrión por su sendero particular en el espacio del desarrollo? La respuesta tiene que ver con el contexto siempre cambiante que se genera con la formación de patrones, pues cada etapa no solo depende de lo que ocurrió antes, sino que también cambia el contexto y, por lo tanto, influye en las etapas siguientes. Esto ocurre porque los genes reguladores pueden responder a otras proteínas gracias a su región reguladora, pero también modifican la actividad de otros genes al sintetizar sus propias proteínas reguladoras. Como las proteínas reguladoras de un núcleo o de una célula son capaces de influir en sus vecinos mediante la señalización molecular, este proceso está coordinado en el espacio y en el tiempo. Las células responden según el contexto particular y al mismo tiempo lo modifican, lo que impulsa al embrión a seguir desarrollándose y le obliga a forjar un camino específico en el espacio del desarrollo. Se trata del principio de la recurrencia, con la constante redefinición del contexto por la aplicación una y otra vez del mismo proceso subyacente.
Si cada etapa del desarrollo embrionario es una modificación de su contexto anterior, entonces podríamos preguntarnos razonablemente de dónde procede el primer contexto. En el caso del embrión de la mosca, comenzamos nuestra historia con un gradiente de la proteína Bicoid, simbolizada por un gradiente de manzanas desde la cabeza a la cola. ¿Cómo surge este gradiente? La respuesta tiene que ver con la mosca madre: el huevo no se forma de forma aislada en su interior, sino conectado por un extremo a células que le suministran diversos componentes, entre ellos copias del ARN del gen Bicoid (recordemos que el ARN es una molécula intermedia que se necesita para fabricar una proteína). Estas copias del ARN están ancladas inicialmente en el extremo de la cabeza del huevo, donde se utilizan para sintetizar la proteína Bicoid (manzanas). La proteína Bicoid y su ARN se propagan entonces por el huevo, formando un gradiente desde la cabeza a la cola[34]. Por lo tanto, el gradiente de manzanas al comienzo del embrión se remonta a un contexto anterior: el huevo y las células que lo rodean en la madre. Al remontarnos más en el contexto del embrión, nos encontramos con el contexto de la madre.
Pero podemos preguntarnos qué es lo que forma el patrón del huevo y de las células que lo rodean en la madre, y encontraremos que esto depende a su vez de cómo se desarrolló la mosca madre. El punto de partida del desarrollo de un organismo también se puede ver como un resultado del desarrollo de la generación anterior. En vez de tener un comienzo claro, nuestro periplo por el espacio del desarrollo forma un ciclo que proporciona a cada adulto el contexto para el periplo de su descendencia. Al igual que la historia narrada en La Odisea, se trata de un viaje de vuelta a sus orígenes, y por eso se le suele denominar el problema del huevo y la gallina (¿qué fue primero: el huevo o la gallina?), que en realidad solo se convierte en un problema cuando queremos definir el primer punto de partida. Es como decir que un círculo es un problema porque no podemos definir un comienzo o un final. Pero en realidad no es un problema; es la naturaleza del círculo. De igual forma, el camino que sigue el embrión por el espacio del desarrollo forma un bucle, de manera que cuando terminamos de recorrer el bucle estamos ya en el siguiente. La cuestión clave aquí no es buscar un comienzo preciso, sino comprender cómo se consigue llevar al embrión por su camino. Tal y como hemos visto, este sendero del desarrollo implica la aplicación recurrente del mismo conjunto de principios subyacentes, en el que el embrión se ve empujado continuamente por su contexto al mismo tiempo que lo va modificando. Gracias a este proceso, el embrión se va transformando desde un inicio muy simple en un individuo pluricelular y complejo. Hemos visto que la recurrencia puede conducir a la elaboración de patrones dentro de un embrión. Pero ¿cómo podemos describir los patrones a muchas escalas diferentes, desde la organización global del cuerpo hasta la organización concreta de las células del ojo o de una hoja? Para responder esta pregunta tenemos que estudiar más a fondo algo que acontece en un segundo plano durante nuestro periplo por el desarrollo: el crecimiento.
§. El lienzo en expansión
George Stubbs estaba obsesionado con los caballos[35]. Nació en Liverpool en 1724 y se formó inicialmente como retratista, pero su interés fue desplazándose poco a poco de las personas a los caballos. Adquirió una reputación sin igual que le permitió conseguir muchos encargos para pintarlos junto sus amos, o juntos en una cacería o en el hipódromo. Por eso no es extraño que cuando en 1762 llegó a Inglaterra un exótico caballo a rayas procedente del Cabo de Buena Esperanza, Stubbs se pusiera rápidamente manos a la obra (figura 27). El retrato de Stubbs constituye la reproducción más exacta y detallada de una cebra en su época, aunque el animal parezca un poco fuera de lugar en un bosque inglés.Para representarla de una forma tan precisa, Stubbs probablemente utilizó numerosos pinceles. Quizá comenzó con pinceles relativamente gruesos que definieran la distribución global del cuadro y luego utilizó pinceles cada vez más finos para introducir mayor detalle. Al variar de esta manera el tamaño de los pinceles, consiguió capturar diferentes escalas de organización, desde la composición general a los detalles minúsculos. ¿Cómo podemos comparar este proceso de perfeccionamiento de las pinceladas con lo que ocurre durante el desarrollo de una cebra real?
Existen varias especies de cebras con un número diferente de rayas. La cebra común (Equus burchelli) tiene unas 25 rayas, la cebra de montaña (E. zebra) unas 40 rayas y la cebra de Grevy o real ( E. grevyi) cerca de 80 (figura 28). (A juzgar por el número y disposición de las rayas del animal, Stubbs pintó una cebra de montaña). Jonathan Bard ha propuesto que la variación entre las diferentes especies de cebras está relacionada con el momento en el que acaece la formación de patrones en el embrión[36]. Los patrones en los que se fundamentan las rayas se cree que aparecen por primera vez en los embriones de cebra de unas pocas semanas, como se muestra a la izquierda de la figura 28. Bard propuso que, en todas las especies, estas rayas iniciales se separan a intervalos regulares de unos 0,4 mm desde el centro de una raya al centro de la siguiente.

Figura 27. Cebra, George Stubbs, 1763.
Este ejemplo resalta la interacción entre la formación de patrones de desarrollo y el crecimiento. A medida que avanza el desarrollo, el número de células se incrementa rápidamente, lo que a menudo está asociado a un incremento de tamaño. Sería como si un lienzo estuviera expandiéndose continuamente mientras el artista trabaja en él. Aunque esto puede sonar más complicado que pintar en un lienzo fijo, hasta cierto punto hace las cosas más simples, puesto que en vez de necesitar una serie de pinceles cada vez más finos para incorporar más detalles, se puede utilizar el mismo pincel todo el tiempo: el mismo pincel que proporciona distinciones generales cuando el lienzo es pequeño, permitirá representar detalles mucho más finos cuando el lienzo es grande. Esto es exactamente lo que ocurre en el ejemplo del desarrollo de la cebra: en el momento en el que aparecen las rayas por primera vez, siempre están espaciadas unos 0,4 mm, pero como el tamaño del embrión va variando, si las rayas aparecen más tarde, cabrán más a lo largo del mismo y darán lugar a subdivisiones más delgadas en el animal adulto.
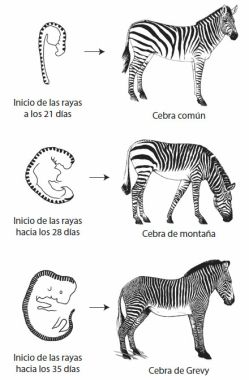
Figura 28. Generación de las rayas en las diferentes especies de cebra.
Para conocer mejor esta interacción entre el crecimiento y la formación de patrones, imaginemos que estamos en el núcleo de un embrión en desarrollo, intentando decidir si encender o apagar determinados genes. Esta decisión siempre es relacional y se basa en las conversaciones locales con las células y núcleos que están más cerca. En las primeras etapas del desarrollo, cuando solo hay unas pocas células, las células más cercanas conformarán la mayor parte del embrión, por lo que cualquier célula recibirá información sobre lo que está ocurriendo en él en su conjunto con bastante eficacia. A partir de esta información, cada célula sabrá si se encuentra en el extremo de la cabeza, y entonces encenderá los genes apropiados. Pero a medida que prosigue el desarrollo y aparecen más células, las cercanas no ocupan más que una fracción cada vez más pequeña del embrión, de manera que la visión del conjunto cada vez resulta más difícil. En consecuencia, cada célula embrionaria va tomando decisiones cada vez con más detalle. Al haber decidido previamente que estaba en el extremo del embrión donde se formará la cabeza, ahora intentará decidir en qué parte de la cabeza se encuentra. Quizá resuelva que se encuentra cerca del frontal de la cabeza, donde se formarán los ojos, por lo que encenderá el siguiente conjunto de genes, que son los adecuados para esta posición. En cada etapa, la célula se compromete con una decisión y la pone en práctica mediante interacciones locales. Como el número de células aumenta continuamente mientras se forman los patrones en los embriones, no hacen falta más mecanismos para explicar el incremento del nivel de detalle que va emergiendo.
§. Deformación
Ya hemos tratado muchos de los principios clave del desarrollo, pero todavía queda otra cuestión que debemos abordar para completar el cuadro. El desarrollo necesita algo más que la mera formación de patrones y un cambio de escala. El adulto no es simplemente una versión magnificada del huevo, en cuyo caso no seríamos más que enormes esferas dando tumbos. No es así: a medida que se desarrollan, las plantas y los animales expanden o contraen con preferencia determinadas posiciones en direcciones concretas. Por ejemplo, una hoja emerge del lado de un tallo, y una pierna nace de un cuerpo. Así que nuestro lienzo no crece de manera uniforme, sino que se expande o se contrae de un modo particular.Para hacerse una idea de lo que esto significa, observemos la figura 29 (lámina 5). En la parte superior se muestra una fotografía de Manzanas y galletas, de Cézanne, que representa algunas manzanas rojas (gris oscuro en la fotografía en blanco y negro) y amarillas (pálido) sobre una mesa. Imaginemos que este lienzo está hecho de un material especial que crece a una velocidad determinada. Con el tiempo, crece más y más rápidamente según las reglas del interés compuesto. Algunos colores, como el amarillo, también tienen la propiedad mágica de que incrementan la tasa local del crecimiento. La parte inferior de la figura 29 ilustra el resultado cuando se simula por ordenador el crecimiento de un lienzo de este tipo. Podemos observar que se ha incrementado su tamaño global y que también se ha deformado por el crecimiento desproporcionado de las manzanas amarillas (pálidas).
Obsérvese, no obstante, que la manzana amarilla de la izquierda se hizo más grande que las otras manzanas amarillas. ¿Cuál es el motivo? La respuesta tiene que ver con el refuerzo y la competencia. Las regiones amarillas del lienzo intentaron crecer más que las otras. Abandonadas a su propia suerte, las manzanas amarillas se harían más y más grandes, con lo que reforzarían continuamente su ventaja de tamaño por la ley del interés compuesto. Pero como no están solas sino conectadas al resto del lienzo, si una manzana amarilla se hace más grande y está rodeada por áreas que no son amarillas, intentará invadir y estrechar sus regiones colindantes, forzándolas a hacerse más grandes para acomodar su tamaño cada vez mayor. Sin embargo, las regiones colindantes se resisten a este estrechamiento ya que intentan crecer a su propio ritmo. Este conflicto entre la manzana amarilla y su entorno crea fuerzas o tensiones en el lienzo que contrarrestan el crecimiento de la manzana amarilla. Se produce esencialmente una competencia por el espacio entre la manzana amarilla y sus vecinos. El resultado es un compromiso en el que las manzanas amarillas crecen menos de lo que les gustaría, mientras que las regiones cercanas se expanden más de lo que querrían. La manzana amarilla de la izquierda está menos constreñida por el material de lento crecimiento porque se encuentra cerca del borde del lienzo, por lo que crece a un ritmo más cercano al suyo y se agranda más que las otras manzanas amarillas.
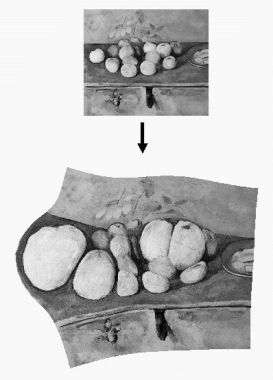
Figura 29. Manzanas y galletas, Paul Cézanne, hacia 1880, y una versión deformada ocasionada por el mayor crecimiento de las regiones amarillas. Véase la lámina 5.
Al introducir el crecimiento diferencial en el lienzo, hemos provocado una nueva forma de refuerzo y competencia. El refuerzo surge porque las regiones que crecen más rápido que otras refuerzan su propio tamaño según la ley del interés compuesto. Pero este proceso está contrarrestado por las tensiones que se generan con las regiones vecinas, lo que conduce a la competencia por el espacio. El resultado global es que el lienzo se deforma de distintas formas.
En nuestra deformación de la obra de Cézanne, los colores del cuadro final modificaron su crecimiento. Ahora imaginemos que ocurre lo mismo mientras se está pintando. Una pincelada de amarillo haría que una región creciera con más rapidez; una mancha de rojo aplicada después sobre el amarillo podría hacer que creciera más despacio. Las regiones estarían continuamente en un tira y afloja de diversas maneras a medida que el cuadro progresa, con lo que se reforzarían constantemente al mismo tiempo que competirían por el espacio. Además de crear un patrón de colores, tales pinturas mágicas acabarían deformando el lienzo de todas las maneras posibles. Este proceso a su vez modificaría el contexto del lienzo, su distribución de colores en el espacio, lo que a su vez influiría en las siguientes etapas del proceso.
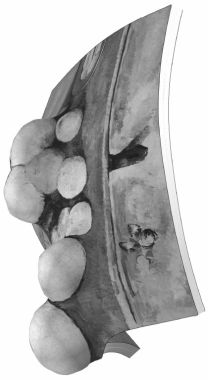
Figura 30. La versión deformada de Manzanas y galletas de Cézanne cuando se deja que el lienzo se combe en tres dimensiones. Véase la lámina 6
El crecimiento de la flor de la boca de dragón (Antirrhinum) ilustra muy bien el surgimiento de las deformaciones por la acción de los genes. A la izquierda de la figura 31 hay un diagrama simplificado de esta flor en una de las primeras etapas del desarrollo, cuando la yema tiene una anchura de aproximadamente 1 mm. En esta etapa, los pétalos forman un pequeño tubo cilíndrico coronado con cinco lóbulos. Imaginémonos que esta es la forma del lienzo al comienzo de un cuadro, con diferentes regiones representadas por sus cinco segmentos originados por algunos de los patrones de actividad génica que se han formado en esta etapa. Por ejemplo, las proteínas reguladoras denominadas CYC y DICH se sintetizan específicamente en los dos segmentos de color más oscuro. En un ordenador podemos utilizar las proteínas reguladoras de la yema de la flor como pinturas mágicas que influyen en el crecimiento del lienzo. A medida que cada región intenta crecer de un modo concreto, el ordenador simula el tira y afloja con las otras regiones que acaba deformando el lienzo[37]. Las pinturas influyen no solo en el ritmo del crecimiento, sino también en la dirección en la que crecen. Por ejemplo, una gradación del color permitiría que el crecimiento se orientara con preferencia en la dirección del gradiente.
El resultado de estas diferentes interacciones se ilustra con la flor de la boca de dragón del centro de la figura 31, que es realmente mucho mayor que el lienzo de partida de la izquierda (su área es unas 500 veces mayor), pero se ha dibujado a un tamaño razonable para facilitar la comparación. La forma final de la flor surge de la interacción entre los genes y de su influencia sobre las propiedades de crecimiento local del lienzo. No es solamente que la forma final se parezca a una boca de dragón, sino que su patrón de crecimiento coincide con la manera en que se sabe que crecen y se desarrollan las diferentes regiones de la flor. Si retiráramos algunas de las proteínas reguladoras de la yema en desarrollo, como las de la región negra, crecería una flor con una forma diferente, como se muestra a la derecha en la figura 31, que se parece a una forma mutante de la boca de dragón que no puede fabricar las proteínas reguladoras CYC ni DICH.

Figura 31. Simulación por ordenador del crecimiento de una flor normal (flor de la izquierda) y una mutante (flor de la derecha) de boca de dragón.
La muerte celular tiene un efecto global parecido en los animales, en los que el desarrollo del sistema nervioso proporciona un buen ejemplo. En este caso, la preocupación no es absorber la luz, sino que las neuronas alcancen las células diana que deben inervar. Durante el desarrollo, nuestro sistema nervioso prolifera para producir más neuronas de las que se necesitarán en el estado adulto. Según la región del cerebro, del 20 al 80% de las neuronas que se producen acabarán degenerándose. Que una neurona sobreviva o no dependerá de la recepción de determinadas moléculas señalizadoras denominadas neurotrofinas[39], que son factores de supervivencia que ayudan a mantener las neuronas vivas. Sin un suministro adecuado de neurotrofinas, una neurona se moriría (mediante un proceso denominado apoptosis). Las neurotrofinas las sintetizan las células diana de las neuronas, de manera que si la neurona se las ingenia para ponerse en contacto eficazmente con una célula diana, recibe una avalancha de neurotrofinas que le permitirá sobrevivir. Pero si no llega a establecerse un buen contacto entre ellas, la neurona probablemente muera. Las neuronas compiten por las neurotrofinas igual que los vástagos de un árbol compiten por la luz, y el resultado global de esta competencia es la poda de algunas neuronas para que solo queden las que formaron buenos contactos con su diana.

Figura 32. Árbol generado por un programa de ordenador basándose en el refuerzo de la ramificación y en la competencia por la luz.
Figura 33. Simulación por ordenador de los cambios morfológicos de Dictyostelium por reorganizaciones y señalización celulares.
§. El lienzo tridimensional
Un lienzo de muchos colores en deformación y en crecimiento ofrece una perspectiva global de lo que ocurre durante el desarrollo. Pero el lienzo es bidimensional, mientras que la estructura de las plantas y de los animales es tridimensional. George Stubbs ilustró la anatomía tridimensional de los caballos al dibujar su estructura interna. Suspendió un cadáver de caballo en una granja y despegó varias capas del animal, mes a mes, dibujando primero los músculos exteriores (figura 34) y finalmente el esqueleto. Fue un trabajo concienzudo realizado seguramente en medio de un hedor insoportable. Stubbs publicó sus resultados en un libro monumental, Anatomía del caballo, la mejor descripción anatómica del animal en su época[40].Estas anatomías complejas surgen a partir de los diferentes procesos que ya hemos explicado. Sin embargo, a partir de ahora tenemos que imaginarlas en un lienzo tridimensional en vez de bidimensional. Para hacernos una idea de lo que suponen estas deformaciones tridimensionales, observemos la figura 35, en la que se muestran algunas de las regiones clave de un embrión típico de vertebrado. En esta etapa, el huevo inicialmente esférico ya se ha transformado en un embrión curvado y tridimensional. Tal y como vimos para la mosca en desarrollo, el embrión aparece descrito con una profusión de temas y variaciones indicados por la regiones sombreadas de diferente forma. Veamos qué ocurre en algunas de estas regiones.

Figura 34. Dibujo de la Anatomía del caballo, George Stubbs, 1766.
Otra estructura clave que sigue a lo largo del embrión es un tubo hueco y largo denominado tubo neural (figura 35), cuya pared está formada por una sola capa de células. Al igual que las somitas, el tubo neural se subdivide a lo largo del embrión en regiones distintas. La mayor parte del tubo neural que discurre por la espalda prolifera para formar distintos elementos de la médula espinal y del sistema nervioso. La región del tubo neural que está cerca de la cabeza forma el cerebro, que crece y se hincha para formar un par de hemisferios cuya pared tiene al principio una sola célula de grosor; a medida que prosiguen el crecimiento y la división celular, las células comienzan a subirse unas sobre otras para formar la corteza cerebral, una capa cuyo espesor tiene varios estratos de células. La corteza en desarrollo también divide su superficie en regiones que expresan diferentes proteínas reguladoras. Por ejemplo, una proteína reguladora está presente en mayor cantidad en el extremo posterior y central de la corteza (figura 36, izquierda[10]). Por el contrario, otra proteína reguladora se expresa más hacia el lateral y el extremo frontal (figura 36, centro). Estas proteínas reguladoras, junto con las moléculas señalizadoras asociadas, conducen a que en la corteza se formen regiones con distintas funciones, tales como las implicadas en el movimiento, la sensibilidad, la audición y la visión (figura 36, derecha). Los ratones que carecen de una o más de estas proteínas reguladoras desarrollan cerebros en los que ha cambiado el tamaño relativo de estas regiones.
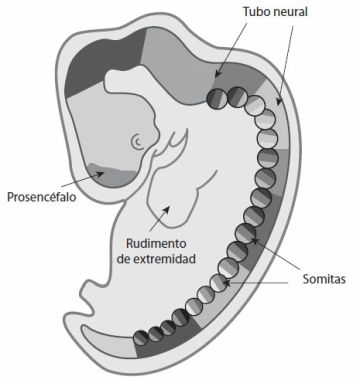
Figura 35. Esquema del embrión de un vertebrado.
§. Una receta común
En el capítulo anterior vimos que el desarrollo se fundamenta en un doble bucle de realimentación entre el refuerzo y la competencia. Las moléculas, como las proteínas reguladoras, refuerzan su propia abundancia o actividad al mismo tiempo que ocasionan su propia inhibición. Este bucle de realimentación está impulsado por colisiones entre un gran número de moléculas (variabilidad de la población), equilibrado por cierto grado de estabilidad molecular (persistencia). En este capítulo hemos visto que algunos otros ingredientes desempeñan una función clave, como la interacción y la cooperación de las moléculas o de las células más cercanas. Esta cooperación entre numerosos componentes introduce un enorme abanico de posibilidades combinatorias, un inmenso espacio de desarrollo de combinaciones moleculares y celulares. Los embriones viven una odisea por este inmenso espacio mediante la aplicación recurrente de nuestro doble bucle de realimentación, que modifica constantemente su propio contexto y, por lo tanto, compone la escena que los lleva a la siguiente etapa del periplo. Mediante este proceso, el embrión crece, se deforma y se forman patrones a distintas escalas.
Figura 36. Corteza de cerebro en desarrollo, en la que se muestran las áreas con proteínas reguladoras clave (izquierda y centro), y las regiones funcionales resultantes implicadas en el movimiento (M), la sensibilidad corporal (S), la audición (A) y la visión (V) (derecha).
Ahora podemos ver que en la evolución y en el desarrollo interviene una interacción parecida entre nuestros siete principios (variabilidad de la población, persistencia, refuerzo, competencia, cooperación, riqueza combinatoria y recurrencia). La evolución y el desarrollo son manifestaciones diferentes de lo que he llamado la receta creativa para la vida. En la evolución, la receta implica poblaciones de individuos en un entorno, y conduce a la aparición de diferentes formas de vida durante muchas generaciones. En el desarrollo, la receta implica poblaciones de moléculas y de células dentro del mismo individuo, y conduce a la aparición de un adulto en una sola generación. Los procesos son muy diferentes, pero podemos reconocer que detrás de esta apariencia diferente subyace una forma común.
Por supuesto, he presentado la evolución y el desarrollo de un modo que resalta los puntos de encuentro. La razón por la cual he seguido esta estrategia es porque nos ayuda a apreciar la esencia de cada proceso. Si consideramos la evolución y el desarrollo por separado, podríamos definir una colección de principios en cada caso, pero probablemente no llegaríamos a la formulación que describo. De hecho, ninguno de estos procesos se suele explicar mediante mi interpretación de la interacción de mis siete principios. Pero si tomamos perspectiva y establecemos comparaciones entre estos procesos, conseguimos conocer lo que es general y lo que es particular de cada caso, lo que nos ayudará a comprender las bases de cómo se producen. En capítulos posteriores veremos que esta misma estrategia nos ayudará a observar otras transformaciones, como el aprendizaje y el cambio cultural.
A pesar de su similitud básica en la forma, también hemos observado una diferencia notable entre la evolución y el desarrollo. La trayectoria que sigue un embrión por el espacio del desarrollo forma un bucle repetitivo, y los embriones van pasando básicamente por las mismas regiones del espacio en cada generación. Esto constituye una gran diferencia con la evolución, en la que las trayectorias que toman las especies por el espacio genético son más indefinidas y no presentan un carácter cíclico. ¿De dónde viene esta diferencia? Para responder esta pregunta necesitamos conocer cómo aparecen en primer lugar las trayectorias del desarrollo. Dejaremos las relaciones de forma para estudiar las relaciones de la historia.Contenido:
§. Comienzos unicelularesBashford Dean tenía dos pasiones en la vida[41]: una era estudiar el desarrollo y la evolución de los peces, gracias a lo cual llegó a ser en 1904 profesor de zoología de vertebrados en la Universidad de Columbia, a la edad de 37 años. La otra pasión era una fascinación por las armas y las armaduras desde su infancia, cuando vio un hermoso yelmo europeo en casa de un amigo de la familia. Lo dejó tan impresionado que se puso inmediatamente a estudiarlo por dentro y por fuera durante mucho tiempo, mientras estaba sentado en el porche de su amigo. Su interés por las armaduras aumentó con los años, y en 1906 se convirtió en conservador honorario de armas y armaduras del Museo Metropolitano de Arte de Nueva York. Finalmente, consiguió jubilarse de sus trabajos de zoología en Columbia para dedicarse en cuerpo y alma a hacer de la colección de armas y armaduras del Museo Metropolitano una de las más excelentes del mundo.
§. El ascenso por la escala
§. Aumentos y crecimiento
§. Una receta dentro de otra receta
Al transferir su interés de los peces a las armaduras, Bashford Dean llevaba a cuestas su pasado biológico, como ilustran una serie de diagramas donde describió la historia de diferentes armamentos, como los yelmos o los escudos, de la misma forma que cualquier otro ilustraría la evolución de los organismos (figura 37). Su diagrama de los yelmos muestra en la parte baja un casco redondo ancestral y simple. A partir de esta forma primitiva emergen diferentes linajes, algunos conducen al yelmo cerrado muy elaborado con visera, y otros conducen a diferentes innovaciones, callejones sin salida o reversiones a formas más simples.
Figura 37. Diagrama de la evolución de los yelmos, Bashford Dean, 1915.
Podemos observar dos procesos entrelazados en la historia de los yelmos. Uno es su cambio del diseño a lo largo de los siglos, la evolución que describe Bashford Dean. El otro es la fabricación, que se repite a sí misma muchas veces cada generación a medida que el fabricante de yelmos va haciendo más copias del mismo. Estos dos procesos están conectados: las acciones del fabricante de yelmos dependen del momento histórico, mientras que la historia de los yelmos depende de cómo los fabricantes modificaron su forma de hacerlos con el tiempo.
Las mismas consideraciones valen para la evolución de los organismos pluricelulares. Aunque solemos retratar la evolución como un árbol ramificado en el que un tipo de organismo parece transformarse directamente en otro, no son los propios organismos los que cambian, sino el modo en el que se desarrollan a partir de los huevos fertilizados. Los primeros productos del desarrollo pueden haber sido entes relativamente toscos, simples amalgamas de células (equivalentes a los cascos más antiguos). Estas estructuras pluricelulares simples surgieron por primera vez hace unos mil millones de años o más. Entonces se desarrollaron las formas más elaboradas del desarrollo que finalmente condujeron a la diversidad de organismos que vemos hoy en día. En cada especie pluricelular viva observamos un proceso de desarrollo parecido que se repite a sí mismo en cada generación para producir continuamente un gran número de organismos de esencialmente la misma clase. Aun así, este proceso repetitivo no está completamente fijado, sino que cambia con el tiempo, mediante la evolución. El desarrollo y la evolución están estrechamente conectados en la historia. La trayectoria del desarrollo depende del pasado evolutivo, mientras que la trayectoria de la evolución depende de la manera en que el desarrollo se modifica con el tiempo.
En los capítulos anteriores hemos visto que la evolución y el desarrollo se basan en la misma receta creativa, y que comparten una forma similar. En este capítulo quiero estudiar otro tipo de relación: la conexión de la historia de la evolución y del desarrollo. Como primera etapa, veamos de dónde vienen los ingredientes básicos para la receta del desarrollo.
§. Comienzos unicelulares
El desarrollo implica la transformación de una célula en un individuo pluricelular; sin embargo, muchos de los principios que lo gobiernan se encuentran ya en los parientes unicelulares[42]. Si pensamos en el comportamiento estadístico de las moléculas, veremos que un organismo unicelular es capaz de metabolizar las moléculas de su entorno, o de importarlas, gracias al movimiento continuo y caótico inherente a cada una de las moléculas. El metabolismo regular y predecible depende de procesos estadísticos, como la difusión, en los que interviene una gran colección de sucesos moleculares. Sin el proceso físico de difusión no existiría ninguna clase de vida. Los organismos unicelulares sacan un gran provecho de la difusión: cuando dos células de levadura de distinto tipo sexual tienen que juntarse para la reproducción sexual, cada una produce moléculas señalizadoras denominadas feromonas que salen de la célula y pasan al entorno, donde se difunden para estimular a las células del tipo sexual opuesto. Como la difusión implica una caída de la concentración con la distancia, proporciona a los organismos información espacial sobre lo cerca o lejos que está la fuente de la feromona: si la concentración de la feromona pertinente es elevada, lo más probable es que una célula del otro tipo sexual esté cerca. La reproducción sexual no es más que un ejemplo de las muchas situaciones en las que los organismos unicelulares utilizan las moléculas señalizadoras y la difusión para comunicarse. El principio de la variabilidad de la población aplicado al desarrollo no es algo que los organismos pluricelulares hayan inventado de la nada, sino que ya había sido explotado por los ancestros unicelulares hace mucho tiempo. Lo mismo ocurre con el principio de la persistencia aplicado al desarrollo, pues demasiada difusión traerá problemas a un organismo unicelular: si una célula no estuviera rodeada por una membrana que evite que las moléculas difundan por el entorno, dejaría de existir como individuo. La persistencia también opera de una generación a otra: a medida que un organismo unicelular se divide, su ADN se copia y el contenido de la célula se comparte entre las células hijas. El contenido incluye a las proteínas reguladoras, lo que permite que las células hijas recreen el patrón de la actividad génica y de las proteínas de su progenitora. Se trata de algo parecido al modo en el que los patrones de la actividad génica se transmiten entre las células a medida que se divide el huevo fecundado de un organismo pluricelular. El mismo proceso de división celular que proporciona persistencia hereditaria entre las generaciones de organismos unicelulares ayuda a mantener la continuidad y la persistencia entre las células dentro del mismo organismo durante el desarrollo.En el mundo unicelular también tenemos los mismos antecedentes sobre los principios de refuerzo, competencia, cooperación y riqueza combinatoria tal y como se aplicaron al desarrollo. Todos los organismos unicelulares regulan sus genes en respuesta a los cambios del entorno. Cuando bebemos un vaso de leche, una bacteria que vive en el intestino, llamada Escherichia coli, modifica la actividad de sus genes para producir más cantidad de una proteína necesaria para digerir la lactosa, o sea, se enciende un gen que codifica la proteína con la que se digiere esta molécula porque la gran cantidad de lactosa de la leche modifica las proteínas reguladoras de la bacteria. Algunas de estas proteínas reguladoras activan genes, mientras que otras los inhiben. Además, al igual que los genes de los organismos pluricelulares, los de los unicelulares tienen regiones reguladoras a las que se pueden fijar muchas proteínas. La intensidad del encendido o del apagado depende del modo en el que interaccionan y cooperan estas diferentes proteínas reguladoras, con lo que se forma un gran número de combinaciones reguladoras diferentes que permiten que la célula responda a los desafíos de su entorno de muchas maneras diferentes.
Del mismo modo que gran parte de las herramientas necesarias para fabricar los yelmos estaban disponibles antes de que aparecieran los primeros, muchos de los elementos necesarios para el desarrollo ya estaban presentes en los organismos unicelulares antes de que evolucionaran los organismos pluricelulares. ¿Cómo se reunieron los diferentes ingredientes en estas formas unicelulares para proporcionar la receta para el desarrollo? Aunque no anduviéramos por ahí para ser testigos directos de estos acontecimientos, podemos hacernos una idea de lo que ocurrió.
§. El ascenso por la escala
La mayor parte del océano está en perpetua oscuridad, pero cerca de la superficie hay suficiente luz para mantener una comunidad próspera de plantas microscópicas[43], algas unicelulares que forman un inmenso prado de plancton marino. Sin embargo, este modo de vida tiene sus peligros: el plancton se verá arrastrado sin remedio por las corrientes marinas y sus turbulencias. Si las plantas microscópicas se ven empujadas a gran profundidad, como la cantidad de luz cae rápidamente, morirán en la inmensa oscuridad, a menos que sepan nadar o flotar para volver a la superficie. Más cerca de la costa hay poca profundidad, lo que les impide caer en la penumbra. Pero no hay garantía de que una planta que viva cerca de la costa no acabe arrastrada hacia el inmenso océano, a menos que se sujete a una roca. Algunas especies de algas unicelulares se agarran al lecho marino, lo que les permite vivir permanentemente cerca de la costa. Estas células suelen tener dos extremos diferentes: uno está especializado en la sujeción a la roca y el otro en la recogida de la energía solar. Con esta disposición incorporan un aspecto de su entorno: la interfase entre el agua marina líquida y transparente, y una roca sólida y opaca.Con este modo de vida conseguirán más luz los individuos que sean capaces de extenderse hacia arriba y de crecer alejándose de la roca. Sus opciones de supervivencia serán mejores que las de sus vecinos ensombrecidos y, por lo tanto, se verán favorecidos por la selección natural. Podría crecer de este modo si sus propias células se hicieran más grandes, pero un organismo unicelular no puede superar ciertos límites de tamaño: a medida que este aumenta, se vuelve más difícil la coordinación de los procesos en el gran volumen de citoplasma. Una solución parcial a esta dificultad consiste en incluir muchos núcleos en la célula, que es lo que suelen hacer las criaturas unicelulares más grandes. Aun así, la mezcla continua del citoplasma dificulta la formación de patrones de actividad génica.
Otra solución a estos problemas consiste en que las células se peguen unas a otras después de la división, lo que permitiría que el organismo se convierta en un individuo pluricelular. Con varias células, cada una con su propio núcleo, surge la posibilidad de encender genes diferentes en células diferentes. Las células cercanas a la roca podrían encender los genes que permiten el agarre, mientras que las que están en el extremo expuesto al agua podrían encender los genes implicados en la fotosíntesis. La evolución de tales células especializadas no requiere un conjunto completamente nuevo de mecanismos puesto que podría surgir de la combinación de ingredientes que ya estaban presentes en los organismos unicelulares. Los principios que se aplican a las células aisladas se aplican a los conjuntos de células que permanecen en proximidad. Entonces, la interfase entre la roca y el mar podría abordarse de otra manera, mediante diferencias entre las células en vez de dentro de cada una. Al organizarse por tipos de células, el alga ha capturado o incorporado una peculiaridad espacial de su entorno, la distinción entre la roca y el mar que la rodea: ha ordenado el mundo al ordenarse a sí misma.
Nuestra alga primitiva es capaz de organizarse siguiendo un sendero tortuoso relativamente simple en el espacio del desarrollo. La célula inicial del alga comienza con una combinación particular de proteínas reguladoras que se corresponden con una posición del espacio del desarrollo. Este contexto proporciona las condiciones moleculares que conducen a la división de la célula y a la producción de unos pocos tipos de células más en las primeras etapas del embrión, algunas mejor adaptadas para agarrarse a la roca y otras para captar luz. El embrión ha sido impulsado a una nueva posición en el espacio del desarrollo, lo que a su vez proporciona el contexto que lo impulsa a la siguiente etapa. Finalmente, las células también se preparan para la formación de las células reproductoras que dan lugar a la siguiente generación. De este modo, el alga se labra un sendero recurrente en el espacio del desarrollo, gobernado por un conjunto de principios moleculares y celulares que ya estaban en funcionamiento en sus ancestros unicelulares.
Esta situación hipotética demuestra que los ingredientes que ya estaban presentes en el mundo unicelular pueden haberse reunido durante la evolución para proporcionar una receta básica para el desarrollo. Una vez que esta receta estaba en marcha, pudieron evolucionar formas más elaboradas de desarrollo. Nuestras algas pluricelulares simples pegadas a la roca compiten por la luz, por lo que las formas capaces de proliferar y hacerse más altas durante su ciclo de vida se verían favorecidas por la selección natural. Al ser más altas, aparecerán otros problemas, como la tensión provocada en la planta cuando se tuerce por las corrientes, o el problema de mantener vivas las células cercanas a la roca a medida que se van alejando de las células fotosintéticas. Puede resultar útil la especialización de otras células, como un collar de células más fuertes para impedir que la planta se desprenda por las corrientes, o un sistema de transporte para mover los azúcares desde el ápice a la base de la planta. No se necesita ningún mecanismo fundamentalmente nuevo para que evolucionen tales especializaciones, puesto que surgirían simplemente por la repetición del mismo proceso de formación de patrones a medida que el organismo se desarrolla. La construcción recurrente de patrones sobre patrones a medida que crece el organismo permite que emerjan de forma organizada un abanico de especializaciones. El resultado global es que se ha extendido y modificado nuestro sendero recurrente por el espacio del desarrollo.
He ofrecido un escenario simplificado para ilustrar la posible integración de nuestros siete principios aplicados al desarrollo durante la evolución de las plantas pluricelulares. Se puede contar una historia parecida con los animales. Muchos animales unicelulares viven felizmente consumiendo a otros organismos. Pero los animales que se hacen más grandes contarán con algunas ventajas, como la capacidad para comerse a otras criaturas y a su vez evitar ser comido[44]]. El incremento de tamaño por volverse pluricelular tiene el beneficio añadido de permitir que se especialicen las diferentes células: algunas se podrían dedicar a comer y otras a digerir la comida. Como los animales que viven en una escala superior se encuentran con otros desafíos, tal como desplazarse de un modo eficaz o coordinar las diferentes partes del cuerpo, podrían surgir más tipos celulares y organizaciones mediante la repetición de la formación de patrones que se ocupan de estos desafíos.
El incremento de tamaño y de complejidad no está exento de ciertos costes. Por ejemplo, retrasa la reproducción porque el organismo necesita más tiempo para crecer hasta su forma madura. Por lo tanto, los beneficios del tamaño necesitan contraponerse a los costes del incremento del tiempo de generación. Los compromisos como este abundan en el mundo de los seres vivos porque la mejora en una dirección suele producirse a expensas de un peor crecimiento en otra. Por esta razón, los organismos pluricelulares no han reemplazado a los unicelulares: estos últimos, como las bacterias, continúan siendo mucho más numerosos que sus parientes pluricelulares. Tal y como Stephen Jay Gould ha apuntado, todavía vivimos en la «Edad de las bacterias», un periodo que ha durado unos 3.500 millones de años[45]. En vez de que la vida progrese de forma global hacia tamaños y complejidades cada vez mayores, en los ecosistemas coexisten muchas formas diferentes, y cada una captura relaciones a diferentes escalas, desde la microscópica a la macroscópica.
§. Aumentos y crecimiento
Cuando miramos un alga marina de hoy en día, por ejemplo un sargazo vejigoso (Fucus vesiculosus, figura 38), vemos que se ha adaptado a su entorno a muchas escalas. Visto el organismo en conjunto, se bifurca y ramifica a lo largo de decenas de centímetros, una adaptación que le permite recoger una gran cantidad de energía solar. En la base produce una estructura que se parece a una raíz y que la ancla a la roca. Este rizoide se conecta al resto de la planta mediante un fuerte tallo que resiste la continua sacudida del mar. Si miramos un poco más de cerca, vemos que el cuerpo principal del alga está aplanado como una hoja, lo que proporciona una gran superficie para recoger energía luminosa. El nervio principal de la hoja le da una fuerza adicional para resistir los movimientos del mar. Algunas regiones también están hinchadas para formar vesículas de aire que le proporcionan flotabilidad. Si observamos la planta con más aumentos, vemos las células, cada una de una décima de milímetro. Hay varios tipos que se especializan de diferentes maneras, como las de la hoja principal, para llevar a cabo la fotosíntesis, o las del rizoide, para adherir la planta a la roca.
Figura 38. Sargazo vejigoso (Fucus vesiculosus).
Todos estos niveles de organización representan todas las escalas en las que el alga marina ha capturado su relación con el entorno. La arquitectura ramificada de la planta se debe a relaciones tales como el modo en el que la luz solar incide sobre la planta, y los efectos de las corrientes marinas y de la gravedad. La forma de la proteína Rubisco se ajusta a las relaciones espaciales de una molécula de dióxido de carbono, es decir, a la disposición de los átomos y de las cargas eléctricas por la superficie de la molécula. A cada escala, las adaptaciones de una planta implican la captura de relaciones espaciales y temporales del entorno con el cual interacciona.
Lo mismo ocurre con los organismos unicelulares. La forma de la proteína Rubisco del sargazo vejigoso es similar a la de las algas unicelulares, y en ambos casos funciona a escala molecular para fijar el dióxido de carbono. Pero algunos aspectos del entorno están ausentes en los organismos unicelulares, como por ejemplo los retorcimientos provocados por las corrientes marinas. Cuando medimos una fracción de milímetro nos podemos agarrar con facilidad a una roca, sin importar lo poderosas que sean las olas que rompen a nuestro alrededor. A esta escala, el agua parecería viscosa, lo mismo que la melaza lo parece a la nuestra. Por lo tanto, incluso aunque la vida unicelular sea rica y variada, hace caso omiso de algunas de las relaciones del mundo a gran escala. Los organismos pluricelulares más grandes tienen que funcionar a esta escala mayor y consiguen capturar muchas de las relaciones que se establecen en este gracias a su diversidad de tipos de células y a la organización a varias escalas. Alcanzan esta organización recorriendo las escalas en sentido opuesto al que antes hemos seguido al acercarnos: mediante el crecimiento y la proliferación. El sargazo vejigoso comienza como un cigoto fecundado que va a la deriva hasta que se ancla en el lecho marino. Entonces se divide en dos células con propiedades diferentes: una de ellas se seguirá dividiendo y generará patrones para formar las frondas de la planta; la otra crecerá, se dividirá y se diferenciará para formar el rizoide. La planta va subiendo por sus distintas escalas de organización mediante la recurrencia del crecimiento y de los patrones.
§. Una receta dentro de otra receta
Durante uno de sus viajes a Europa, el amante de las armaduras Bashford Dean se topó con una caja vieja arrumbada en un ático antiguo. La caja había pertenecido a un armero allá por el año 1600 y contenía partes de guanteletes sin acabar. Bashford Dean recordaba así el episodio:Tenía curiosidad por sentir el tacto de estos objetos antiguos que parecía que su fabricante hubiera puesto ayer en la caja. Tenía la sensación de que si salía por la vieja puerta que tenía al lado, sería objeto de un encantamiento como Alicia en el País de las Maravillas y pasaría al siglo XVI para encontrarme en la otra habitación un verdadero armero en su mesa cerca de la ventana.[46]En este capítulo, también nos hemos transportado al pasado. Hemos visto que la evolución y el desarrollo comparten una conexión histórica. Muchos elementos del desarrollo ya estaban al acecho en los ancestros unicelulares que existieron hace miles de millones de años. Estos ingredientes se acabaron por integrar de un modo concreto durante la evolución de los organismos pluricelulares. Las primeras formas pluricelulares mostraron poca recurrencia durante el desarrollo, por lo que mostraban una organización simple con dos o tres tipos celulares. Estos cambios representaban senderos tortuosos relativamente simples a través del espacio del desarrollo. Pero a medida que fueron apareciendo más niveles de recurrencia y que los senderos se hicieron más complicados y extensos, las criaturas podían explorar y capturar las relaciones con su entorno a muchas escalas, lo que dio lugar a nuevos modos de sobrevivir y reproducirse con eficacia.
En vez de permanecer fijos, los senderos tortuosos del desarrollo están en continua reorientación y deformación en la escala evolutiva del tiempo porque los periplos por el espacio genético y del desarrollo están conectados[47]. A medida que las poblaciones se desplazan por el espacio genético durante la evolución, los genes que influyen sobre las trayectorias del desarrollo pueden cambiar para permitir que los organismos capturen nuevas relaciones de su entorno. Desde esta perspectiva más amplia, el desarrollo no siempre sigue con precisión el mismo sendero ni regresa exactamente al mismo lugar después de una generación: las trayectorias se van desplazando poco a poco, por lo que durante el tiempo evolutivo forman una serie de espirales en vez de bucles estrictos. Si nos remontamos con la gallina y el huevo lo suficientemente atrás en el tiempo, no encontramos ni gallinas ni huevos, sino sus primeros ancestros, organismos que estaban constituidos por tan solo unos pocos tipos de células. Si vamos más atrás incluso en el tiempo, no encontraríamos ninguna trayectoria del desarrollo, sino simplemente el ciclo reproductor de los organismos unicelulares.
Al igual que las muñecas rusas, la evolución y el desarrollo muestran una relación doble. Por una parte, el desarrollo está incrustado históricamente en la evolución, pues surgió del proceso evolutivo y está contenido en él. Por otra parte, el desarrollo tiene una forma similar a la de su progenitor evolutivo, pues se basa en la misma receta creativa. Aunque uno opere en muchos individuos y generaciones y el otro en un único individuo y generación, en ambos casos encontramos unos principios fundamentales comunes.
Sin embargo, no hemos llegado al final de la historia. Del mismo modo que la evolución dio origen al desarrollo, veremos que el desarrollo conducirá a otro proceso, el aprendizaje, con la misma receta. Para continuar con esta próxima transición, primero debemos conocer mejor el funcionamiento del aprendizaje. En vez de intentar conocer inmediatamente la manera en que aprenden los seres complejos como los perros o los humanos, buscaremos primero los fundamentos en el comportamiento de los organismos más sencillos.Contenido:
§. Se hacen ajustesUno de los cuadros más curiosos del Renacimiento es una representación meticulosa de un trozo de terreno cubierto de hierbas de Alberto Durero (figura 39). Durero extrae diseño y harmonía de un conjunto aparentemente aleatorio de hierbas al que normalmente no prestaríamos atención. Al elegir un tema tan mundano, es capaz de transmitir su talento artístico de una forma pura, sin contaminar por distracciones convencionales. De igual forma, los científicos a menudo eligen estudiar temas sencillos cuando intentan llegar a la esencia de un problema. Cuando se estudian los sistemas relativamente simples se evitan complicaciones y distracciones innecesarias, y se consiguen sacar conocimientos más profundos. Esto es particularmente cierto cuando intentamos comprender algo tan problemático como nuestra capacidad para aprender. Las reacciones humanas son tan complejas y están tan entrelazadas con nuestras emociones que puede ser difícil interpretarlas con objetividad. A veces ayuda dar un paso atrás y plantearse cómo se enfrentan a los retos los seres más modestos, como las bacterias, las hierbas o las babosas.
§. El relato de la flora
§. El mordisco de Venus
§. Una babosa de mar sensible
§. Patrones temporales
§. Respuestas humanas
§. La ordenación del mundo
En el capítulo anterior estudiamos la manera en la que los organismos se enfrentan a los retos espaciales, como distinguir entre la roca y el mar. Gracias al desarrollo adquieren distinciones anatómicas que les permiten capturar parte de la heterogeneidad que les rodea. En este capítulo quiero empezar a analizar la dimensión temporal y la manera en que los organismos afrontan el cambio. Si el entorno fuera siempre constante, un organismo nunca tendría que modificar su comportamiento según las circunstancias. En esta situación, los procesos como el aprendizaje serían mucho menos importantes. Para llegar a la raíz del aprendizaje, primero tendremos que conocer las respuestas básicas de los organismos ante los cambios que les rodean. Consideraremos un abanico de respuestas en los microorganismos, en las plantas y en los animales, para así comenzar a discernir los elementos comunes y los distintivos. Por ejemplo, las plantas no tienen neuronas, pero no obstante son capaces de capturar de su entorno patrones de cambio bastante complejos. Una vez que hayamos establecido los mecanismos biológicos básicos para afrontar los cambios, podremos estudiar cómo se integraron estos diferentes mecanismos para dar lugar al aprendizaje, nuestro tercer ejemplo de la receta creativa para la vida.

Figura 39. Porción de césped o Estudio de malas hierbas, Alberto Durero, 1503.
§. Se hacen ajustes
Vivimos en un mundo fluctuante en constante cambio. En vez de ser inflexibles, a los organismos les resulta beneficioso poder ajustarse a los cambios según las circunstancias. Ya nos hemos encontrado una manera de responder a los cambios: como se mencionó en el capítulo anterior, la bacteria Escherichia coli puede ajustar su nivel de actividad génica según la cantidad de lactosa de su entorno. Tiene proteínas reguladoras que encienden o apagan genes en respuesta a la concentración de lactosa. Esta respuesta es una adaptación que permite que la bacteria sintetice proteínas, tal como la enzima que digiere la lactosa, que le ayudarán a sobrevivir y a reproducirse. Todos los organismos unicelulares poseen mecanismos de regulación de esta clase que les permiten responder a muchos factores que podrían cambiar con el tiempo, desde la prevalencia de determinadas moléculas a la temperatura o la luz.En algunos casos, estas respuestas pueden ser casi inmediatas. El alga verde unicelular Chlamydomonas, que suele encontrarse en las charcas, siente la luz y se mueve hacia ella gracias a dos filamentos muy delgados denominados flagelos (figura 40). Las proteínas receptoras sensibles a la luz que están en la célula de Chlamydomonas se modifican cuando la luz incide sobre ellas, y esto conduce a la generación de una señal eléctrica que la recorre rápidamente. La señal cambia el patrón de los movimientos de los flagelos, por lo que la célula se vuelve hacia la luz, al igual que un nadador ajusta su brazada para girar. Al nadar hacia la luz, Chlamydomonas se asegura de que puede permanecer en un medio brillante y continuar sobreviviendo mediante la fotosíntesis.
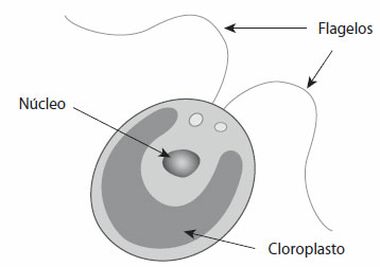
Figura 40. Célula de Chlamydomonas.
Sin embargo, las respuestas instintivas no tienen por qué ser fijas, sino que se pueden ir ajustando con el tiempo. Un buen ejemplo es el comportamiento nadador de la bacteria Escherichia coli[48]. Al igual que Chlamydomonas, Escherichia coli nada con su flagelo, aunque en este caso el flagelo actúa como propulsor rotatorio en vez de por flexión. La bacteria nada instintivamente hacia las áreas ricas en sustancias alimenticias, como los azúcares, y se aleja de los compuestos nocivos como los ácidos. Lo hace gracias a las moléculas receptoras que hay en la membrana, que perciben continuamente la concentración de las sustancias químicas que la rodean. Estos receptores influyen en el movimiento de la bacteria mediante una estrategia similar a la de un niño jugando a «frío, frío»: los movimientos se ajustan según la bacteria se aleja (frío, frío) de la fuente de interés o se acerca (caliente, caliente) a ella. La bacteria comienza su marcha en una dirección aleatoria, y si percibe que las condiciones van mejorando (que aumenta la concentración de azúcares), entonces continúa en la misma dirección. Pero si nota que las condiciones van a peor, se da la vuelta y nada en otra dirección seleccionada prácticamente al azar. Después de nadar y girar unas pocas veces, acabará finalmente por alcanzar la fuente de comida.
La bacteria Escherichia coli se comporta así debido a una propiedad notable de sus receptores: su sensibilidad se modifica gradualmente según el nivel de estimulación. Si hay mucho azúcar alrededor de la bacteria, sus receptores se van haciendo poco a poco menos sensibles al azúcar, por lo que cuesta más estimularlos. Esto significa que si el azúcar abunda durante mucho tiempo, la bacteria se habitúa y ya no responderá a la concentración elevada del mismo. La bacteria solo responde con fuerza cuando cambia la concentración de la comida. Si está nadando hacia una fuente de azúcar, percibe el incremento de la concentración y mantiene la marcha; si está alejándose de dicha fuente, nota que la concentración disminuye y cambia de dirección. La bacteria no sólo responde a la situación predominante, sino que compara su percepción con la historia reciente de las percepciones para determinar si la disponibilidad de comida mejora o no. Captura las relaciones con el tiempo y las utiliza para determinar su camino en el espacio.
La selección natural ha hecho evolucionar muchas respuestas instintivas como adaptaciones a entornos variantes. Estas respuestas capturan las relaciones espaciales y temporales del entorno. Cuando alguien bebe un vaso de leche, esta avanza por el espacio hasta llegar al intestino, donde inunda el entorno de la bacteria Escherichia coli con un aporte de lactosa. A medida que la Tierra rota en el espacio, cambiará el modo en que la luz incide sobre un charco y también lo hará la dirección en la que Chlamydomonas nadará. A medida que Escherichia coli se va desplazando, podría toparse con una fuente rica de alimento y nadar hacia ella. El tiempo y el espacio están conectados y muchos elementos de esta relación se capturan gracias a las respuestas instintivas.
Dado su predominio en los organismos unicelulares que viven hoy en día, las respuestas instintivas deben haber surgido muy pronto en la evolución biológica, cuando el mundo de los seres vivos comprendía solo organismos unicelulares. Pero con la llegada de los organismos pluricelulares complejos hace unos 500 millones de años, estas respuestas se llevaron a una escala diferente: en lugar de que las células reaccionen por separado, las respuestas se coordinan entre muchas células de un individuo. El mundo de las plantas proporciona algunos ejemplos sorprendentes.
§. El relato de la flora
Si el cuadro de Durero de un trozo de césped es una de las curiosidades del arte renacentista, entonces los cuadros de Giuseppe Arcimboldo conseguirían el premio a la excentricidad. En El bibliotecario (figura 41, lámina 7) utiliza libros y papel para construir un hombre barbudo completo con un brazo y una mano. La yuxtaposición inusual de los objetos y significados confiere a este cuadro más afinidad con los cuadros surrealistas del siglo XX que con otras obras renacentistas. Pero quizá el ejemplo más famoso del trabajo de Arcimboldo es su descripción de las cuatro estaciones (figura 42, lámina 8), donde compone cabezas humanas a partir de trozos de plantas de las distintas épocas del año. En Primavera utiliza flores para componer la cabeza; en Verano utiliza frutas; en Otoño monta la cabeza con productos de recolección tardía, como las patatas y las uvas; mientras que para Invierno utiliza un tocón nudoso. Las plantas proporcionan el material ideal para ilustrar las estaciones porque sufren cambios importantes en el transcurso del año.
Figura 41. El bibliotecario, Giuseppe Arcimboldo, 1565. Véase la lámina 7.

Figura 42. «Las cuatro estaciones»: Primavera (arriba a la izquierda), Verano (arriba a la derecha), Otoño (abajo a la izquierda) e Invierno (abajo a la derecha), Giuseppe Arcimboldo, 1573. Véase la lámina 8.
La capacidad para percibir la luz es común en las plantas. La luz es una parte importante de su existencia porque proporciona la entrada de energía que resulta esencial para la fotosíntesis, por lo que las plantas han desarrollado numerosas formas de percibir la cantidad y la calidad de la luz. Lo consiguen mediante una serie de proteínas receptoras que ponen en marcha una serie de reacciones cuando incide la luz, como son el encendido y apagado de genes. Por lo tanto, el primer requisito para medir la duración del día (la capacidad para detectar y responder a la luz) no solo se encuentra en los animales, sino que también constituye una característica básica de los sistemas vegetales.
Quizá lo más sorprendente es que las plantas también tienen un reloj interno[49]. Lo descubrió por primera vez el astrónomo francés Jean-Jacques d’Ortous de Mairan en 1729, al querer conocer el motivo por el que las mimosas pliegan su hojas por la noche y las abren durante el día. Para ver si estaba relacionado con la variación de la cantidad de luz a lo largo del día, colocó algunas en la oscuridad dentro de un armario. Ojeándolas de vez en cuando se dio cuenta de que las plantas seguían abriendo y cerrando las hojas con el ritmo diario incluso aunque se mantuvieran continuamente en la oscuridad. La variación de la intensidad lumínica externa no resultaba esencial para mantener el ritmo, sino que parecía deberse a cierta clase de proceso interno, como si la planta tuviera un reloj.
Hace tan solo unas décadas conseguimos conocer el funcionamiento de los relojes biológicos en las plantas y en los animales. En el caso de las plantas, surgió gracias al estudio de Arabidopsis thaliana, una mala hierba que no hubiera estado fuera de lugar en el césped de Durero. Es el equivalente vegetal de la mosca de la fruta, con una capacidad similar para reproducirse con rapidez en el laboratorio. Los estudios con Arabidopsis revelaron que su reloj interno se basa en el aumento y la disminución diarios de determinadas proteínas reguladoras[50]. Recordemos que las proteínas reguladoras se fijan a los genes y los activan o inhiben. Las que intervienen en el reloj suben y bajan su concentración cada 24 horas como resultado de un circuito de realimentación. A medida que aumenta en una célula la concentración de una de las proteínas reguladoras activadoras, comienza a encender la síntesis de una proteína inhibidora. Entonces, la proteína inhibidora comienza a incrementarse e impide que se sintetice más activador. Si no hay más activador, el inhibidor ya no se produce y la concentración de la proteína activadora comienza a incrementarse de nuevo, con lo que se completa el circuito. Se tardan unas 24 horas en realizar un ciclo completo de este tipo. El sistema de realimentación es similar a lo que ocurre en el modelo de Turing descrito en el capítulo 3, salvo que aquí estamos creando patrones temporales y no espaciales.
Hay otra característica clave en el reloj vegetal: suele ponerse en hora mediante sucesos externos como la salida del sol, como si se ajustara un reloj a una hora fija cada día al amanecer. Esto no altera el ritmo del reloj porque sigue tardando 24 horas en realizar un ciclo. El ciclo se suele desplazar para coincidir con las fluctuaciones ambientales diarias, lo que no ocurre cuando las plantas se mantienen artificialmente en la oscuridad continua, con lo que el reloj continúa funcionando sin tener en cuenta el entorno. El desplazamiento se produce porque las transiciones entre la oscuridad y la luz influyen en el ritmo cíclico de las proteínas reguladoras. La aparición de la luz matinal conduce a un cambio molecular en el ajuste del reloj que persiste al menos hasta la siguiente salida del sol, cuando el reloj se vuelve a poner en hora. Por consiguiente, el reloj de la planta no programa los acontecimientos de una manera fija, sino que lo hace en función de sucesos externos como la salida del sol.
Regresemos ahora al problema de percibir la duración del día. Las plantas lo consiguen mediante la integración de su capacidad para percibir la cantidad de luz con la información de su reloj interno. A medida que los días se alargan en la primavera, las hojas detectan la luz cada vez más tarde con respecto a la salida del sol. Estos retrasos corresponden a las fases más tardías del ciclo interno de las proteínas reguladoras. Finalmente, la detección de la luz en una fase particularmente tardía del ciclo de la proteína hace que las hojas sinteticen una molécula señalizadora que viaja hasta los ápices en crecimiento de la planta. A continuación, la molécula activa los genes que promueven el desarrollo de las flores, con lo que comienzan a formarse los primordios florales. Las flores de primavera que vemos cada año dependen de esta integración entre la percepción de la luz, el ajuste de la hora y la señalización. Gracias a este proceso, muchas plantas consiguen detectar la separación temporal relativa de dos sucesos, la salida y la puesta del sol y, por lo tanto, determinan el momento del año.
Al ajustarse a las estaciones, las plantas consiguen capturar las relaciones temporales, que a su vez están enclavadas en el espacio. El ajuste del reloj de la planta está relacionado con la rotación diaria de la Tierra sobre su eje, y los cambios estacionales de la duración del día son consecuencia de la rotación anual de la Tierra alrededor del Sol. Las plantas coordinan esencialmente su desarrollo con los movimientos planetarios, no porque tengan algún interés por la astronomía, sino porque se benefician de florecer en determinados momentos del año: florecer en el momento apropiado es una respuesta instintiva que permite que las plantas se reproduzcan con más eficacia.
Este relato muestra que la floración de las plantas responde a peculiaridades temporales y espaciales de su entorno, incluso aunque estén formadas por muchos millones de células. La respuesta de cada célula está coordinada por señales que viajan entre ellas, por lo que la planta consigue responder como un todo. He descrito la respuesta de las plantas a la duración del día, pero no es más que una de las muchas respuestas a su entorno. Las plantas también integran su reacción a factores como la temperatura y los nutrientes. Cuando entran en floración, están tomando una decisión basada esencialmente en un enorme abanico de factores ambientales, y lo mismo ocurre con otras respuestas, tales como la caída de las hojas o la germinación de la semilla.
Estas respuestas provocan cambios en una planta que perdurarán horas o meses. La aparición algo más temprana del sol por la mañana alterará el reloj interno de proteínas reguladoras, alteración que durará al menos hasta la siguiente salida del sol. Basta con que la planta se exponga a uno o dos días más largos para que inicie la formación de las flores, y una vez que empiece la floración, continuará durante muchas semanas, incluso si la planta se vuelve a poner en condiciones de días cortos. Los días largos condujeron a un cambio permanente en la actividad génica, lo que a su vez condujo a una modificación duradera de la organización espacial mediante la producción de flores.
Gracias a la persistencia de los cambios, las plantas son capaces de responder a una secuencia de sucesos temporales en vez de tan solo sucesos únicos y aislados. El amanecer desplaza el reloj interno de la planta y, por lo tanto, modifica su respuesta a la luz el resto del día. Un acontecimiento matinal dura hasta el atardecer, lo que permite que una respuesta se base en otra, y permite que una planta detecte si es un día de primavera o de invierno. Esto ocurre porque las plantas no solo responden a su entorno, sino que algunas respuestas pueden cambiar la capacidad de respuesta de la propia planta. Esto se parece a la capacidad de respuesta de la bacteria nadadora Escherichia coli, que ajusta su sensibilidad según la cantidad de azúcar que acaba de percibir a su alrededor.
Aunque pueda impresionarnos lo elaborados que son los mecanismos de las respuestas instintivas de las plantas (como el inicio de la floración), nos parecerán lentos porque sus efectos pueden tardar semanas o incluso meses en resultar evidentes. Por eso tendemos a verlos como parte del ciclo anual y no como reacciones sofisticadas. Nos sorprenden más las respuestas que ocurren a una escala de tiempo más corta. Las reacciones visibles y rápidas son más raras en las plantas que en los animales, pero existen algunos ejemplos informativos.
§. El mordisco de Venus
Si un insecto merodea por una hoja de la Venus atrapamoscas (Dionaea muscipula), la hoja se cierra de golpe[51] (figura 43). El infortunado insecto queda pues atrapado por una jaula de «dientes» entrelazados en el borde de la hoja y va siendo digerido lentamente por las secreciones de la superficie interna de misma, lo que permite que la planta recoja nutrientes muy valiosos. Charles Darwin estaba fascinado por la Venus atrapamoscas, y la denominó «la planta más maravillosa del mundo»[52]. Había observado que la respuesta de cierre foliar está activada por unos pelos sensibles al tacto en su superficie interna que se estimulan cuando se doblan por el movimiento del insecto. Una vez estimulados, las células de los tricomas generan un pulso eléctrico que se detectó por primera vez en 1873 gracias a John Burdon-Sanderson, colaborador de Darwin. Este pulso, conocido como un potencial de acción, se expande por la hoja y lo perciben sus células, lo que ocasiona que se cierre de golpe por un mecanismo hidráulico. La respuesta táctil de la Venus atrapamoscas es muy veloz (0,3 s) para enfrentarse a los movimientos rápidos de su presa.
Figura 43. La hoja de la Venus atrapamoscas en sus formas abierta (izquierda) y cerrada (derecha).
La Venus atrapamoscas ha desarrollado estructuras que le permiten capturar las relaciones espaciales y temporales. A partir de la secuencia de estímulos puede decirse cuándo es probable que un insecto merodee por una hoja. La respuesta también tiene especificidad espacial: solo se cerrará la hoja estimulada. La organización espacial de la planta (esto es, las muchas hojas que ha generado durante el desarrollo) le permite responder según la posición. Esencialmente, es capaz de saber dónde y cuándo se posó un insecto.
En el caso de la Venus atrapamoscas, los estímulos se refuerzan mutuamente para incrementar una respuesta, pero también hay casos en los que la repetición de la estimulación tiene el efecto opuesto y reduce la respuesta. Si tocamos los folíolos de Mimosa pudica, se plegarán rápidamente entre sí, con lo que se cerrará la hoja (figura 44) gracias a una señal eléctrica desencadenada por el movimiento de los folíolos cuando se produce un contacto. En 1960, cuando un tifón azotó Tokio, la investigadora Hideo Toriyama de la Universidad de Mujeres notó que después de un tiempo de contactos continuos, las plantas locales deMimosa pudica dejaban de cerrar sus hojas y las mantenían abiertas[53]. Era como si las hojas se hubieran acostumbrado al fuerte viento y entonces ignoraran la estimulación continua. Toriyama fue capaz de repetir este efecto más tarde en condiciones controladas al colocar las plantas frente a un ventilador eléctrico. Demostró que necesitaba unas diez horas para que apareciera la habituación. En este caso, las respuestas persistieron y se sumaron para reducir la sensibilidad global, igual que vimos para la habituación de Escherichia coli a la concentración elevada de azúcar.
Figura 44. Hojas de Mimosa púdica antes (izquierda) y después (derecha) de la estimulación.
Todas estas respuestas dependen de la organización espacial de las plantas en un intervalo de escalas. En el control del momento de la floración intervienen partes de la planta como las hojas, los tallos y los ápices en crecimiento. La reacción de la Venus atrapamoscas depende de los tricomas sensibles al tacto incluidos en cada hoja. Como vimos en los capítulos anteriores, la organización anatómica surge durante el desarrollo: el óvulo fecundado de la planta crece y se divide para formar raíces, tallos y hojas, con sus numerosos tipos de células. Estos patrones de desarrollo son adaptaciones que permiten que la planta sobreviva mejor en un entorno que también presenta patrones espaciales y temporales.
También vemos que las respuestas al entorno se realimentan para alterar el desarrollo. El desarrollo de las flores puede depender de una respuesta a un factor ambiental, como la duración del día. Tendemos a pensar que el desarrollo es un proceso fijo que ocurre casi sin tener en cuenta su entorno. Quizá adoptemos este punto de vista porque el comienzo de nuestro propio desarrollo, en el vientre de nuestra madre, está protegido. Pero la separación entre el desarrollo y las respuestas al entorno no son tan nítidas: muchas respuestas instintivas conducen a cambios permanentes en la organización corporal, en el patrón de las células y en su conexión con el organismo. Estos cambios pueden producirse mientras el organismo crece y se desarrolla. No existe una división clara entre el desarrollo y las respuestas al entorno, sino que están muy entrelazadas. Nos hemos fijado en esta interacción en las plantas, pero veremos que se aplican principios similares en el caso de los animales.
§. Una babosa de mar sensible
Durante su viaje a bordo del Beagle, a Charles Darwin le cautivaron las reacciones reflejas de una gran babosa de mar que observó cuando estuvo en las Islas de Cabo Verde. En su diario anotó lo siguiente: «Abundan las Aplysia grandes de una longitud de unos 12,5 cm y un color amarillento sucio jaspeado con púrpura… Cuando se le molesta, emite un chorrito de líquido rojo purpúreo, que tiñe unos 30 cm del agua que lo rodea»[54]. Durante mucho tiempo, la reacción de este animal no fue más que una curiosidad zoológica. Pero las cosas cambiaron en los años sesenta, cuando Eric Kandel, en la Universidad de Nueva York, decidió utilizar una variedad californiana de esta babosa, Aplysia californica (figura 45) como modelo para estudiar en detalle los mecanismos de las respuestas animales[55]. Al igual que otros animales, las respuestas reflejas de Aplysia dependen de células nerviosas, también conocidas como neuronas. La ventaja de la babosa sobre muchos otros animales para la investigación es que las neuronas son relativamente grandes, lo que facilita su manipulación. Tampoco hay muchas: el cerebro de Aplysia comprende unas 20.000 neuronas, mientras que nuestro cerebro ronda los 100.000 millones o más. Pero antes de entrar en detalles sobre este sistema, examinemos algunas características generales de neuronas.
Figura 45. La babosa de mar Aplysia californica.
Cada neurona puede estar en reposo o activada. Durante la mayor parte del tiempo, una neurona estará en reposo sin que ocurra gran cosa. Sin embargo, una neurona puede pasar al estado activado, durante el cual el cuerpo celular enviará un pulso eléctrico, o potencial de acción, por el axón. El efecto sobre la siguiente neurona dependerá de lo que ocurra cuando el pulso llegue en las sinapsis del extremo del axón, pues cada sinapsis actúa como un repetidor que convierte la señal eléctrica entrante en una señal química que atraviesa la separación entre una célula y la siguiente. Cuando un impulso eléctrico llega a una sinapsis, se liberan las moléculas señalizadoras, denominadas neurotransmisores, y rápidamente se difunden por la separación para estimular los receptores de la membrana celular del otro lado. Los neurotransmisores tienen la capacidad de excitar o inhibir. En el caso de excitar, incrementan la posibilidad de que se active la célula del otro lado de la separación, mientras que en el caso inhibidor, el neurotransmisor disminuye la posibilidad de activación de la célula siguiente. Según el tipo de neurotransmisor liberado y de los receptores del otro lado de la separación, una sinapsis puede ser estimuladora o inhibidora.
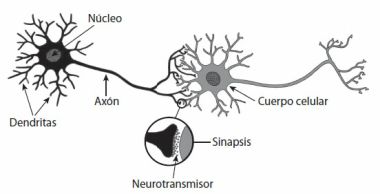
Figura 46. Dos neuronas.
El reflejo de retirada de las branquias es solo una de las tantas maneras de responder a los cambios de su entorno que tiene la babosa de mar. La figura 48 muestra la organización de su sistema nervioso central completo. Hay nueve aglomerados o ganglios, y cada uno contiene varios cientos de células nerviosas. Los diferentes tipos de señales sensoriales tienden a viajar a los diferentes ganglios. El reflejo de retirada de las branquias, por ejemplo, implica que los receptores táctiles de la piel envíen sus señales al ganglio abdominal. Las señales procedentes del ojo se dirigen a otros ganglios, los ganglios cerebrales, mientras que las de la boca van a los ganglios bucales.
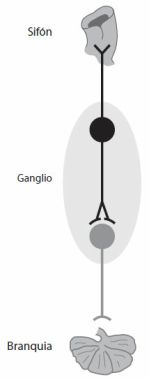
Figura 47. Las neuronas sensoriales (negra) y motoras (gris) conectadas por sinapsis en el ganglio abdominal de la babosa de mar Aplysia californica.
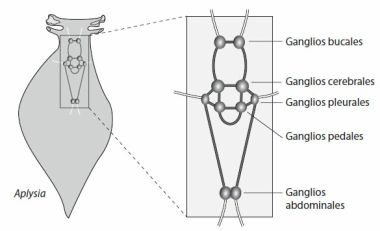
Figura 48. El cerebro de Aplysia.
Sin embargo, los ganglios no funcionan como si estuvieran aislados. Lo que come una babosa de mar puede relacionarse con lo que ve: una pieza sabrosa de alga marina tiene un aspecto diferente a un amasijo de roca insípido porque hay asociaciones características entre la composición de los objetos y el modo en el que reflejan la luz. La babosa de mar es capaz de captar tales relaciones e integrar la información de distintos ganglios, los cerebrales y los bucales en este caso. Es capaz de aprender, por ejemplo, que el aspecto del alga marina es un buen predictor de comida comestible, pero no así una roca (retomaremos cómo ocurre este aprendizaje en el siguiente capítulo). Por lo tanto, discurren entre los ganglios muchos axones que llevan información de uno a otro y que posibilitan las respuestas a asociaciones y distinciones más amplias. De este modo, la anatomía del animal captura las relaciones de su entorno en un amplio intervalo de escalas. Todas estas relaciones anatómicas se establecen en el animal gracias al desarrollo. La organización del sistema nervioso de la babosa de mar, desde el patrón de los ganglios hasta cada neurona sensorial y motora con sus sensibilidades y conexiones iniciales, surge a medida que se desarrolla desde el huevo. La organización interna que se desarrolla entonces permite que la babosa de mar discrimine entre diferentes características de su entorno y responda en consonancia. Al igual que las plantas, la babosa de mar ordena el mundo ordenándose primero a sí misma.
§. Patrones temporales
El sistema nervioso de los animales responde a acontecimientos individuales, pero también puede responder a las secuencias de sucesos en el tiempo. Si estimulamos repetidamente el sifón de la babosa de mar, se debilitará progresivamente el reflejo de retirada de las branquias, del mismo modo que las hojas de la mimosa se habitúan a las sacudidas continuas del viento. Se trata de un caso de lo que Marcel Proust denominaba el efecto anestesiante de la costumbre: bajamos el nivel de respuesta a medida que algo que percibimos como peligroso se va volviendo familiar.En todas las formas de habituación, el efecto del estímulo tiene que persistir durante algún tiempo para que debilite la respuesta siguiente. En el caso de la babosa de mar, esta persistencia tiene dos formas: a corto plazo y a largo plazo. Cuando estimulamos el sifón 40 veces seguidas, provocaremos una habituación que dura aproximadamente un día. Es decir, la babosa de mar tendrá menos capacidad de respuesta al estímulo táctil del sifón durante un día, para luego regresar a la normalidad. Sin embargo, si estimulamos la babosa de mar diez veces cada día durante cuatro días, la habituación dura semanas, esto es, la reducción de la capacidad de respuesta persistirá durante mucho más tiempo. ¿Qué hay detrás de estos cambios a corto y largo plazo?
La habituación a corto plazo conlleva cambios en la intensidad de las sinapsis en el ganglio (figura 49). La activación continuada de la neurona sensorial (negra) conduce a un debilitamiento de las sinapsis que la conectan a la neurona motora (gris), y reduce la eficacia de la señal, lo que se representa por un menor tamaño de las conexiones sinápticas en el diagrama central de la figura 49. El debilitamiento de las sinapsis implica un cambio molecular en las terminaciones de la neurona negra: después de la llegada de un impulso eléctrico, liberarán menos cantidad de glutamato, el neurotransmisor de excitación. Después de un tiempo, cada sinapsis vuelve a su intensidad normal, lo que explica la corta duración del cambio de sensibilidad.
La habituación a largo plazo da lugar a un cambio más duradero en las sinapsis, que no solo se debilitan permanentemente, sino que también hay menos: cambia la anatomía además de cambiar el funcionamiento de cada sinapsis. Esta circunstancia se muestra en el diagrama de la derecha de la figura 49, donde una única sinapsis conecta las neuronas gris y negra. En estos cambios más duraderos en las sinapsis intervienen genes que se encienden y se apagan en el núcleo de la neurona sensorial negra. La estimulación repetida de una sinapsis da lugar inicialmente a un debilitamiento temporal, al igual que con la habituación a corto plazo. Esto marca, en cierto modo, que la sinapsis ha cambiado. Si se repite el patrón de estimulación durante días, las moléculas de señalización se envían una y otra vez desde la sinapsis debilitada al núcleo de la neurona sensorial negra, lo que hace que determinados genes se enciendan y se apaguen, y finalmente conduce a un debilitamiento más permanente de la sinapsis marcada, y algunas veces a su eliminación. Al igual que con la floración de las plantas, la regulación génica interviene en la persistencia a más largo plazo.
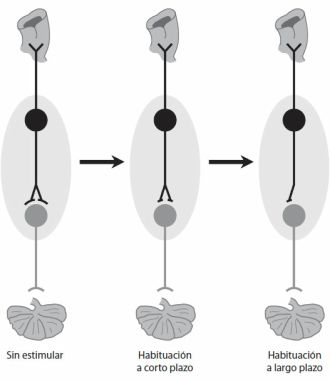
Figura 49. Efectos de la habituación a corto y largo plazo.
El mecanismo de sensibilización es complementario de muchas maneras al de la habituación. En la sensibilización a corto plazo, se refuerzan temporalmente determinadas sinapsis, lo que corresponde a un incremento de la liberación de neurotransmisores cuando llega un impulso eléctrico. En la sensibilización a largo plazo, no solo se refuerza la permanencia de las sinapsis, sino que se crean la mayoría de ellas, y se incrementa el número de contactos entre las células nerviosas. Por ejemplo, en vez de dos sinapsis entre las células nerviosas negra y gris, el número podría incrementarse a cuatro o cinco. Al igual que en la habituación a largo plazo, interviene la activación de genes por las señales emitidas entre las sinapsis y el núcleo.
§. Respuestas humanas
El cerebro humano es mucho más elaborado que los ganglios de la cabeza de la babosa de mar, pero comparte algunas características básicas de organización. Al igual que en esta, la principal función del cerebro humano consiste en integrar la información sensorial y generar las respuestas apropiadas. Pensemos en cómo respondemos al tacto: nuestras neuronas cutáneas sensibles al tacto tienen axones que se extienden hasta la médula espinal, una columna larga de tejido neural alojado dentro de las vértebras (figura 50). Para los reflejos simples, la médula espinal actúa como el punto de encuentro para las neuronas sensoriales y motoras, una función similar a la desempeñada por el ganglio abdominal de la babosa de mar. En el reflejo rotuliano, al golpear la rodilla se activan las neuronas sensoriales y estas envían una señal por su axón hacia la médula espinal, donde las terminaciones axónicas están conectadas mediante sinapsis con las neuronas motoras, que repiten la señal de vuelta a la rodilla y activan la contracción muscular y el movimiento de la pierna.El cerebro solo interviene cuando la respuesta es más complicada y requiere un mayor nivel de integración. Cuando cogemos una manzana, notamos el tamaño y la forma del fruto en nuestra mano, lo que, a su vez, influye en la forma de agarrarla. Sin el flujo de información desde la mano al cerebro y viceversa, quizá la agarremos con poca fuerza y se caiga. Esto es mucho más complicado que una simple respuesta refleja. Cuando agarramos la manzana, las señales sensoriales de la mano llegan a la médula espinal y se transmiten a través de otras neuronas hasta el cerebro, donde se integran las señales de muchas áreas diferentes. Una de estas es una franja de la capa externa del cerebro, o corteza, que recibe el nombre de área somatosensorial primaria (figura 51, arriba).

Figura 50. El sistema nervioso humano.
Aunque el mapa somatosensorial sea una distorsión, el orden de sus regiones refleja en buena parte la del cuerpo. La región que corresponde a la pierna es la siguiente a la del pie. De igual forma, las regiones que corresponden a los dedos de la misma mano están cerca unas de otras. Al igual que en la babosa de mar, esta disposición espacial refleja las correlaciones que existen en el patrón de las entradas: cuando sostenemos una manzana, los receptores del tacto repartidos por la mano se estimulan juntos, mientras los otros no se ven afectados. Si metemos el pie en la bañera, los receptores de las piernas y de los pies se activan juntos, mientras que otros permanecen mudos. Nuestra organización neural es el reflejo de las relaciones que encontramos con más frecuencia a nuestro alrededor.
Figura 51. Áreas somatosensorial y motora de la corteza humana mostrada en perspectiva lateral (arriba) y como secciones transversales (centro y abajo).
El principal propósito de integrar toda esta información sensorial consiste en responder con las acciones apropiadas. Al ver una manzana, podemos extender el brazo y recogerla con la mano. Luego podemos apretar o aflojar el agarre según la información que recibimos de la mano. En todos estos actos intervienen señales de áreas sensoriales de nuestro cerebro que influyen en las neuronas y modifican lo que hacemos.
Al igual que existen regiones en el cerebro consagradas a la información sensorial, hay regiones que se encargan de nuestras acciones. Un buen ejemplo es el área motora primaria. Penfield encontró que cuando estimulaba esta área, que se encuentra frente al área somatosensorial (figura 51), se activaban determinadas articulaciones: se podía contraer un dedo o se podía flexionar un tobillo, porque las señales del área motora estimulada seguían vías determinadas que las conducían a la parte superior de la médula espinal. A continuación iban a la médula y activaban las neuronas motoras, que se excitaban a un determinado nivel, provocando las contracciones musculares específicas. De nuevo, Penfield encontró que las regiones del área motora estaban organizadas de un modo que reflejaba las del cuerpo: las regiones que controlan la mano y los dedos están cerca unas de otras, mientras que las que controlan el pie están cerca de la de los dedos del pie (figura 51, abajo). Esto tiene sentido porque nuestras acciones suelen implicar con mucha frecuencia la coordinación del movimiento de las partes cercanas. Cuando apretamos la manzana, todos los dedos de la mano se mueven juntos, y esto se refleja en la organización de las regiones del cerebro que conducen a estos movimientos.
El sistema nervioso humano está organizado en una serie completa de escalas, desde la arquitectura global de los nervios que van y vienen de la médula espinal, hasta el cerebro con sus diferentes áreas y subregiones. Gran parte de esta complejidad anatómica surge por el desarrollo. En el capítulo 4 vimos que el embrión forma un tubo neural que poco a poco se va subdividiendo y elaborando para producir la médula espinal y el cerebro. Un ser humano es una gama de patrones neurales en marcha que debe su origen al proceso de elaboración que se produjo en el útero.
§. La ordenación del mundo
Todas las respuestas que hemos explicado en este capítulo permiten que un organismo distinga determinados sucesos, o secuencias de sucesos, en su entorno. La bacteria Escherichia coli puede distinguir si está nadando hacia la comida o alejándose de ella, una planta puede distinguir los días cortos de los largos, una Venus atrapamoscas discrimina entre un insecto y una ráfaga de viento, una babosa de mar distingue el toque de un depredador de la sacudida inofensiva del mar, y un humano sabe si su mano agarra una manzana o un trozo de roca. Estas respuestas permiten que los individuos categoricen el mundo, que lo ordenen según determinados sucesos. Estas categorizaciones son posibles porque el mundo no es homogéneo sino que está muy organizado a muchas escalas. La materia se reúne para formar soles, planetas, rocas, mares y seres vivos. El movimiento y las interacciones entre distintos conjuntos de materia conducen a toda una serie de cambios correlacionados. A medida que nuestro planeta rota, la iluminación del sol en muchos puntos de la superficie terrestre sube y baja al mismo tiempo, países enteros se hunden en la oscuridad o vuelven a la luz. Cuando se cae una manzana, todos los átomos que contiene se mueven hacia abajo al unísono. Nuestro entorno es un contexto rico en el que intervienen muchos tipos diferentes de cambios correlacionados.Las respuestas instintivas ayudan a que un organismo discrimine entre algunas de estas correlaciones, y le permiten responder a los sucesos más importantes para su supervivencia y reproducción. La Venus atrapamoscas se cierra de golpe cuando el movimiento colectivo de las moléculas de un insecto estimula los pelos de la hoja, mientras que no los estimulan otros cambios, como el movimiento de las moléculas de aire. La planta disocia un tipo de circunstancia de su entorno (un insecto en movimiento) de otro (aire en movimiento) gracias a que la fuerza ejercida por las moléculas estrechamente asociadas de un insecto es más intensa que la del gas. Al responder a una secuencia de sucesos en vez de a uno, la Venus atrapamoscas distingue además entre un insecto y el impacto de un poco de arena. La misma capacidad discriminatoria aparece en las respuestas animales: tocar una babosa de mar provoca su retracción, mientras que el agua que normalmente circula a su alrededor no tiene ningún efecto. La colisión con las moléculas estrechamente asociadas en un sólido tiene un impacto mayor que un líquido, y es más eficaz para activar las terminaciones sensoriales cutáneas. Al responder a una secuencia de acontecimientos de este tipo, la babosa es además capaz de discriminar entre lo que probablemente resulte inofensivo y lo que no. Hasta el más humilde de los organismos categoriza el mundo espacial y temporalmente.
Pero tal clasificación presenta otro aspecto: la capacidad de los individuos para responder y discriminar entre los sucesos de su entorno depende de sus propias subdivisiones o distinciones internas. La respuesta de una planta a la duración del día está entretejida en su distribución anatómica, en su organización en una serie de hojas, tallos y yemas. La respuesta de una Venus atrapamoscas depende de la disposición espacial de los tricomas sensoriales y de la estructura de la hoja. La respuesta de una babosa de mar o un humano depende de si tiene células nerviosas especializadas con unas sensibilidades y conexiones concretas que se organizan a muchas escalas diferentes como reflejo del abanico de correlaciones que el animal encuentra en su entorno.
En los capítulos anteriores hemos visto que esta organización del individuo va emergiendo durante el desarrollo. La organización interna resultante permite que cada individuo responda de maneras específicas. Pero las respuestas de un organismo también pueden realimentarse para influir en su propia organización. Así, una hierba podría activar la producción de flores en respuesta a la duración del día, y la intensidad de las conexiones neurales de la babosa de mar, o su número, podrían cambiar en respuesta al tacto. En todos estos casos, la respuesta conduce a un cambio persistente o duradero en el organismo. Estos cambios de la forma de organizarse pueden ocurrir mientras el organismo todavía está creciendo y desarrollándose. Una planta produce flores a medida que crece y una babosa responde a su entorno antes de alcanzar la fase adulta. No hay una separación nítida entre el desarrollo y las respuestas que engendra porque la organización interna no está fija, sino que hasta cierto punto puede cambiar en respuesta a su entorno, incluso mientras está en desarrollo. No es ya que nuestras respuestas instintivas sean posibles gracias al desarrollo, es que están entretejidas en el propio desarrollo. Todas las respuestas que hemos abarcado hasta ahora venían dictadas por el pasado evolutivo del organismo. Durante muchas generaciones, determinadas respuestas se han visto favorecidas por la selección natural, como el modo en el que una bacteria responde al azúcar, una planta a la duración del día, o un animal al contacto. Muchos animales también exhiben respuestas a elementos ambientales con los que nunca se habían topado antes en su pasado evolutivo. Un perro puede aprender a reconocer la voz de su amo, un sonido al que los lobos ancestrales nunca se habían expuesto. O una rata puede aprender un camino en un laberinto al que nunca se habían expuesto sus predecesores. En estos procesos intervienen nuevas relaciones entre los elementos de su entorno, como la relación entre un sonido y una persona, o entre una serie de vueltas en un laberinto. Las plantas y los microorganismos no parecen tener esta capacidad para aprender nuevas relaciones. El césped no puede aprender a agacharse cuando comienza a sonar un cortacésped. No se debe a que haya restricciones a la hora de percibir o responder a los sonidos, sino a que las plantas parecen incapaces de aprender relaciones nuevas durante su vida. Incluso aunque las plantas y los microbios sean capaces de responder de maneras muy elaboradas y sofisticadas a su entorno, sus respuestas son siempre instintivas, basándose en la experiencia previa de su pasado evolutivo.
La capacidad de aprender relaciones nuevas es algo peculiar de los animales con sistemas nerviosos elaborados, como las moscas, las babosas, los perros y los humanos. ¿Y cómo ocurre? Como veremos, intervienen muchos de los componentes descritos en este capítulo, pero integrados de un modo particular. Aparece en nuestro tercer caso de la receta creativa para la vida.Contenido:
§. El perro profetaEntre los cuadros más sorprendentes de Miguel Ángel se encuentran las cinco sibilas en el techo de la Capilla Sixtina (figura 52). Las sibilas eran mujeres de la antigüedad con inspiración divina que tenían el poder legendario de ser capaces de ver el futuro. En las pinturas de Miguel Ángel podemos ver que las sibilas sostienen libros especiales en los cuales están escritas las profecías. A las sibilas se les consultaba por lo mismo que se consulta hoy en día a las pitonisas: para conocer lo que se nos avecina. Damos un gran valor a ser capaces de predecir el futuro porque si sabemos lo que está a punto de ocurrir, podemos prepararnos y ajustar nuestras acciones en consecuencia.
§. Neuronas predictivas
§. Se aprende de las discrepancias
§. Pávlov y los castigos
§. Principios fundamentales
§. Un periplo neural
§. Quedarse en marcha
§. Una receta para el aprendizaje
La predicción del futuro no es solo un atributo de sibilas, profetas y pitonisas, sino que forma parte de la vida cotidiana: predecimos tan a menudo que apenas somos conscientes de ello. Cuando volvemos una página de este libro, esperamos encontrarnos otro texto. Si nos encontráramos las páginas en blanco, nos sorprendería y nos preguntaríamos si hubo algún problema en la impresión del libro. De igual forma, si soltamos el libro en algún lugar y nos vamos a tomar un café, esperamos que el libro se encuentre donde lo dejamos a nuestro regreso. Podría parecer más bien pretencioso llamar predicciones a tales expectativas modestas, pero eso es lo que son. Cada expectativa que tenemos sobre el futuro es una predicción, una anticipación de lo que ocurrirá después. Si no estuviéramos haciendo predicciones continuamente, la vida nos resultaría imposible porque estaríamos en un estado continuo de ansiedad, sin conocer nunca ni lo que habrá a la vuelta de la esquina ni lo que debemos esperar[57].

Figura 52. Sibila de Libia, Miguel Ángel Buonarroti, 1511.
La capacidad de predicción de los humanos y de otros animales está muy influida por el aprendizaje. Un recién nacido no tiene las expectativas elaboradas de un adulto porque todavía tiene que adquirirlas aprendiendo de la experiencia. De igual forma, si lanzamos una pelota a un cachorro de perro, ni la mirará ni se anticipará como lo hace un perro más experimentado. La predicciones empiezan a mejorar por el aprendizaje inmediatamente después del parto: un recién nacido aprenderá muy pronto a esperar la comida, y le seguirán rápidamente otras expectativas. No obstante, algunos sucesos son más predecibles que otros. Predecimos con facilidad que el sol saldrá mañana o que la primavera llegará en pocos meses, pero estaremos mucho menos seguros de si mañana lloverá a las tres de la tarde o del número ganador de la lotería, conocimientos que nos resultarían muy valiosos, pero que desafían nuestros poderes de predicción. La magia de las sibilas procede no de su capacidad para predecir, que todos la compartimos, sino de tener poderes proféticos que sobrepasan lo que solemos conseguir.
A diferencia de las sibilas, nosotros, al igual que otros animales, adquirimos la capacidad predictiva no con poderes mágicos ni consultando volúmenes proféticos, sino mediante libros neurales escritos por nuestra experiencia. ¿Y cómo se escriben estos libros? En el capítulo anterior, vimos que nuestras respuestas pueden cambiar por la modificación de las neuronas. La sensibilidad al tacto que tiene la babosa de mar puede incrementarse o disminuirse con la modificación de las sinapsis en las primeras experiencias. Se trata de cambios instintivos en nuestras respuestas, muy constreñidas por el pasado evolutivo. Pero ¿qué hay del aprendizaje de las cosas nuevas? Nuestros ancestros hace 10.000 años no tenían libros, y a los ancestros lobunos de los perros tampoco les tiraban pelotas. Aun así, los humanos y los perros mantienen unas expectativas particulares para cada caso. ¿Cuál es la base neural de este aprendizaje? Al intentar responder a esta pregunta nos introduciremos de lleno en nuestra manera de adquirir el conocimiento acerca del mundo. Tal y como hemos visto en los capítulos anteriores, para abordar las preguntas más complejas nos resultará útil plantearnos primero ejemplos más simples. Un buen punto de partida para los fundamentos neurales del aprendizaje es el perro de Pávlov.
§. El perro profeta
El científico ruso Iván Petróvich Pávlov comenzó sus estudios sobre el aprendizaje en torno a 1900, cuando rondaba los cincuenta años. Por entonces ya había realizado trabajos pioneros sobre el aparato digestivo, durante los cuales había elaborado un procedimiento simple que le permitió medir la cantidad de saliva producida por un perro vivo. Cuando le enseñaba comida al perro, Pávlov podía determinar fácilmente la cantidad de saliva adicional que producía. Observó que se podía desencadenar la salivación del perro con muchos factores, con tal de que estuvieran asociados a la presentación de la comida: «Tan solo con ver el recipiente del cual se le ha dado la comida se consigue evocar un reflejo alimentario completo con todos sus detalles; y, además, la secreción puede estar provocada incluso cuando ve a la persona que trajo el recipiente, o por el sonido de sus pisadas»[58]. Pávlov decidió estudiar esto sistemáticamente. Eligió un estímulo que normalmente no desencadenaría una mayor salivación, como el sonido de una campana; Pávlov hacía sonar la campana cada vez que iba a dar de comer al perro. Con el tiempo, el perro comenzó a salivar más siempre que se hacía sonar la campana, incluso si no se le daba de comer: su respuesta había quedado condicionada por la campana. Tal condicionamiento pavloviano a menudo se interpreta como una demostración de que un perro aprende a asociar el sonido de la campana con la comida. De hecho, suele hacerse referencia al condicionamiento pavloviano como ejemplo del aprendizaje asociativo. Pero se puede interpretar de otra manera: quizá el perro está aprendiendo a predecir, más que aprendiendo una asociación. Por el sonido de la campana es posible predecir que la comida está de camino, y quizá por eso la campana desencadena mayor salivación. ¿Cuál es la diferencia entre la asociación y la predicción?Cuando se para la rueda de la ruleta y la bola cae en una casilla, el croupier anuncia el número. Podemos decir que hay una asociación entre el número que anuncia el croupier y el número de la casilla donde cayó la bola. Los dos van juntos. Pero no se nos ocurriría decir que el croupier predice la casilla en la que acabará la bola porque da el resultado después de que la bola se ha detenido. Si el croupier dijese correctamente el número antes de que la bola se parase, diríamos que la había predicho correctamente. Por lo tanto, nuestra idea de lo que es o no una predicción se basa en la secuencia temporal. En cambio, podemos tratar asociaciones sin ninguna referencia al orden temporal. Por ejemplo, podemos observar una asociación entre el anuncio del croupier y la casilla en donde cayó la bola independientemente de si la bola se para antes o después de que el croupier lo anuncie.
Si el perro de Pávlov está aprendiendo una asociación, entonces no debería importar si la campana suena antes o después de que se presente la comida. Sin embargo, resulta que la hay: si la campana suena justo después, en vez de antes, de darle la comida, la campana no llega a desencadenar una mayor salivación que cuando suena sola. De hecho, el sonido de la campana más bien tiende a tener un efecto inhibidor de la salivación. Parece que el valor predictivo de la campana es lo que importa, y no la simple asociación de la campana con la comida. Si la campana suena antes de la comida, significa que la comida está a punto de aparecer y el perro saliva más preparándose para este acontecimiento. Si la campana suena después de que se haya dado la comida, significa que la comida desaparecerá aunque se la esté comiendo, por lo que el perro se prepara para este acontecimiento salivando menos. El perro está aprendiendo predicciones, no simples asociaciones. La idea de que la respuesta del perro implica una predicción es coherente con otro tipo de experimento, en este caso llevado a cabo por Leon Kamin en Princeton en los años sesenta del siglo XX con ratas, aunque ilustraré los resultados con perros[59]. Supongamos que, en vez de un sonido de campana, condicionamos al perro con otro tipo de estímulo, como tocarle la pata. Si siempre se la tocamos antes de darle la comida, el perro finalmente aprende a salivar más después de tocarle la pata, no es más que la respuesta pavloviana tradicional que cabría esperar. Podemos continuar con este condicionamiento hasta que el perro esté completamente entrenado para responder al toque de la pata. ¿Qué ocurre si le aplicamos repetidamente dos estímulos simultáneos, tocarle la pata y sonar la campana, justo antes de darle la comida? Si el perro aprende a asociar independientemente cada señal con la comida, entonces debería aprender a responder a la campana y al contacto, porque ambos están ocurriendo a la vez antes de comer. Pero el perro no aprende a salivar en respuesta únicamente a la campana, aunque todavía salive en respuesta únicamente al contacto. Esto tiene sentido si el perro está aprendiendo según el valor predictivo del estímulo. Si ha aprendido que el toque en la pata significa que la comida está de camino, la campana no añade más información, ya no tiene valor predictivo, por lo que el perro no aprende a conectarla con la comida. De nuevo, lo importante es la secuencia temporal: si en vez de hacer sonar la campana al mismo tiempo que le tocamos la pata, hacemos sonar la campana antes, entonces el perro aprende a salivar en respuesta a la campana, incluso si se le ha entrenado antes tocándole la pata. La campana ahora está proporcionando la información adicional de que es el primer aviso de que la comida está de camino, por lo que el perro aprende esta nueva relación. El condicionamiento pavloviano sirve para la predicción.
Parece que el cerebro del perro hace profecías basadas en su pasado, aprendiendo de experiencias anteriores para anticipar el futuro. ¿De qué clase de mecanismo dependerá este aprendizaje? Mientras Pávlov realizaba sus estudios en Rusia, otros científicos como Santiago Ramón y Cajal en España y Charles Sherrington en Inglaterra investigaban los detalles del funcionamiento de las neuronas[60]. Demostraron que una característica clave de las neuronas reside en el modo en el que se activan mutuamente mediante las sinapsis. ¿Qué relación tienen estas investigaciones sobre las neuronas con los resultados de Pávlov? Los primeros intentos de responder esta pregunta los hicieron dos científicos a finales de los años cuarenta del siglo XX: el neurofisiólogo polaco Jerzy Konorski y el psicólogo canadiense Donald Hebb. En su libro Conditioned reflexes and neuron organisation («Reflejos condicionados y organización neuronal», 1948), Konorski demostró que los resultados de Pávlov podrían interpretarse por la intervención de cambios específicos en la formación y en el número de conexiones sinápticas entre las neuronas[61]. Donald Hebb llegó de forma independiente a un concepto similar en su libroLa organización de la conducta (1949, traducido en 1985[62] ). A pesar de estos estudios pioneros, quedó poco claro cómo aparecían los cambios sinápticos y cómo podrían conducir a mejorar las predicciones. Tuvieron que transcurrir 40 años más para que se retomara el tema del condicionamiento, cuando se identificaron las neuronas implicadas en el aprendizaje predictivo.
§. Neuronas predictivas
A mediados de los años ochenta del siglo XX, Ranulfo Romo y Wolfram Schultz de la Universidad de Friburgo (Suiza) estaban registrando la señal eléctrica de ciertas neuronas del cerebro de un mono a medida que aprendía tareas[63]. Estas neuronas están localizadas en una región conocida como el mesencéfalo y en sus terminaciones sintetizan un neurotransmisor denominado dopamina. Recordemos que los neurotransmisores son sustancias que se liberan desde las terminaciones axónicas y atraviesan el espacio sináptico para influir en la actividad de la siguiente neurona. La activación de las neuronas liberadoras de dopamina que Romo y Schultz estudiaban parecía estar relacionada con las recompensas que recibía el mono. Se conectó una caja a la jaula del mono que a veces contenía una recompensa, por ejemplo un trozo de manzana. El mono no podía ver el contenido de la caja, por lo que en principio desconocía que pudiera contener algo de comida. Pero si el mono metía la mano en la caja, a veces notaba un trozo de manzana y lo recogía. Romo y Schultz encontraron que las neuronas que estaban registrando se activaban con más frecuencia cuando el mono metía la mano en la caja y encontraba el trozo de manzana (figura 53, panel superior). Primero sospecharon que la activación podría estar relacionada con los movimientos del brazo del mono, pero después demostraron que las neuronas no incrementaban su tasa de activación cuando el mono no encontraba un trozo de manzana en la caja. Al parecer, la activación de las neuronas dopaminérgicas estaba relacionada con el contacto de la manzana, y no con los movimientos del brazo.
Figura 53. Activación de las neuronas dopaminérgicas antes y después del condicionamiento.
Se entenderá mejor si reformulamos el experimento del mono sobre la base de tres conceptos: recompensas, expectativas y discrepancias. Inicialmente, el mono no tenía expectativas de recompensa (el trozo de manzana) cuando escuchaba el sonido de la puerta. Por consiguiente, cuando se encontraba la puerta abierta y cogía un trozo de manzana había una discrepancia entre lo que el mono esperaba (puerta cerrada y sin recompensa) y lo que encontraba (puerta abierta y recompensa). El efecto del aprendizaje consiste en eliminar esta discrepancia: el sonido de la puerta lleva al mono a esperar una recompensa. Sin embargo, el mono todavía no sabe cuándo sonará la puerta porque no tiene manera de saberlo, así que en este punto todavía cabe un elemento de sorpresa o una discrepancia en las expectativas. La discrepancia se ha desplazado en realidad desde el momento en el que el mono recibe la recompensa hasta un momento anterior, cuando suena la puerta. La actividad neuronal que Romo y Schultz estaban midiendo parecía relacionada con la cronología de las discrepancias, con el momento en el que el mono se sorprende. El problema reside en explicar cómo un desplazamiento cronológico de este tipo puede surgir por las interacciones neurales.
§. Se aprende de las discrepancias
Unos años después, los neurocientíficos computacionales Read Montague, Peter Dayan y Terry Sejnowski, en los Estados Unidos, propusieron una solución ingeniosa que integraba los hallazgos experimentales de Romo y Schultz con una teoría matemática del aprendizaje denominada aprendizaje por diferencias temporales (aprendizaje DT[64] ). El aprendizaje DT había sido formulado por Richard Sutton y Andrew Barto varios años antes. Merece la pena sumergirse en el mecanismo propuesto por Montague, Dayan y Sejnowski para conocerlo con más detalle porque sirve para resaltar los principios clave del aprendizaje.En la figura 54 se ilustra una versión simplificada de su esquema, donde se ven dos neuronas que reciben diversos impulsos entrantes. He denominado a la de arriba neurona de expectativas, pero esto no quiere decir que tenga propiedades fisiológicas especiales que le permitan esperar una cosa u otra. Se trata de una neurona como otra cualquiera, con impulsos de entrada y de salida, cuya función en el esquema, como veremos, es la de reflejar las expectativas del mono. La neurona de expectativas recibe información de muchas señales sensoriales (como el sonido de la apertura de la puerta) o estímulos visuales (como un destello). Estas señales están conectadas a la neurona de expectativas mediante sinapsis débiles, que se ilustran con pequeños botones en las terminaciones axónicas. Al principio, estas señales estimulan poco o nada a la neurona de expectativas.
Figura 54. Esquema del aprendizaje DT que muestra la fuerza de las conexiones neurales antes del condicionamiento.
El elemento final del esquema es que la activación de la neurona de discrepancias altera a su vez las señales que le llegan a la neurona de expectativas. Lo hace de un modo más bien especial: la activación intensa de la neurona de discrepancias tiende a reforzar las sinapsis de las neuronas que han sido activadas justo antes de que se activara la neurona de discrepancias. Si la neurona sensible al sonido de la puerta se ha activado antes de la acción de la neurona de discrepancias, entonces la fuerza de su sinapsis se incrementa ligeramente cuando se activa la neurona de discrepancias. Sin embargo, si la neurona del sonido de la puerta no se hubiera activado antes de la activación de la neurona de discrepancias, entonces no se incrementan sus sinapsis con la neurona de expectativas, que podrían incluso perder fuerza.
Ahora estamos en posición de ver cómo funciona el aprendizaje predictivo. Supongamos que el mono toca la manzana después de escuchar la puerta. Percibir la manzana hace que la neurona de discrepancias se active a una tasa elevada (debido a la fuerza de la sinapsis estimuladora). Cualquier impulso que llegue a la neurona de expectativas que se activó justo antes de esto verá ligeramente reforzada su sinapsis con la neurona de expectativas. Esta situación también vale para la neurona del sonido de la puerta, pero no para la neurona del destello. Incluso aunque las neuronas del sonido de la puerta y del destello consigan enviar impulsos a la neurona de expectativas, solo se refuerza la sinapsis con la neurona del sonido de la puerta porque se activa inmediatamente antes que la neurona de discrepancias (que a su vez fue estimulada por la entrega de la recompensa). El refuerzo de una sinapsis dependerá de si se ha estimulado justo antes de que se reciba la recompensa.
Al haberse activado recientemente, la neurona del sonido de la puerta refuerza con eficacia su propia fuerza con la ayuda de la neurona de discrepancias. Este auto refuerzo continúa con otras rondas de apertura de puerta y de retirada de comida, lo que conduce a la formación de una sinapsis cada vez más fuerte entre la neurona del sonido de la puerta y la neurona de expectativas, lo que se indica mediante la mayor longitud de la sinapsis en la figura 55. Podríamos esperar que este auto refuerzo continúe indefinidamente, y que la sinapsis entre ambas neuronas sea cada vez más fuerte. Sin embargo, esto no ocurre porque la neurona del sonido de la puerta finalmente sucumbe ante su propio éxito, como veremos a continuación.
A medida que se incrementa la fuerza de la sinapsis con la neurona del sonido de la puerta, el patrón de activación de la neurona de discrepancias comienza a cambiar de dos maneras. Primero, la neurona de discrepancias comienza a activarse con más intensidad cuando se abre la puerta porque se activa la neurona del sonido de la puerta y estimula a la neurona de expectativas mediante una sinapsis más fuerte. El aumento de la activación de la neurona de expectativas a su vez estimula a la neurona de discrepancias (recordemos que la neurona de expectativas tiene efecto estimulador cuando aumenta su nivel de activación). Por lo tanto, la neurona de discrepancias ahora se activa aún más cuando el mono escucha que se abre la puerta.
En segundo lugar, el patrón de activación de la neurona de discrepancias cambia en el momento de la entrega de la recompensa. Como antes, tocar la manzana estimula la activación de la neurona de discrepancias. Pero a medida que ocurre esto, también hay una caída en el nivel de activación de la neurona de expectativas porque el mono ya no escucha el sonido de la apertura de la puerta. Según la regla descrita antes, esta caída de la activación de la neurona de expectativas conduce a la inhibición de la neurona de discrepancias, que ahora recibe dos impulsos antagonistas: uno estimulador desde la manzana de recompensa, y otro inhibidor por la reducción de la activación de la neurona de expectativas. Este efecto inhibidor de la neurona de expectativas significa que la neurona de discrepancias se activa menos que la primera vez que recibió la recompensa, lo que a su vez significa menos refuerzo de la sinapsis con la neurona del sonido de la puerta, que contrarresta la tendencia de esta neurona a estimularse a sí misma.
Figura 55. Esquema de aprendizaje DT que muestra el refuerzo de las conexiones neurales después del condicionamiento. Al compararla con la figura 54, la sinapsis con la neurona del sonido de la puerta es más fuerte, y el movimiento del brazo del mono ahora está acoplado a la activación de la neurona de discrepancias.
Este esquema del aprendizaje DT explica cómo puede surgir el desplazamiento temporal de la activación de las neuronas dopaminérgicas estudiadas por Romo y Schultz. Estas neuronas se corresponden con la neurona de discrepancias en nuestro esquema. Inicialmente, la neurona de discrepancias se activa a un nivel elevado cuando se percibe la recompensa, pero después del aprendizaje, la subida de la activación se adelanta al momento en el que el mono escucha el sonido de la apertura de la puerta. Este adelanto es exactamente lo que se observa en las neuronas dopaminérgicas, lo que sugiere que se están comportando como las neuronas de discrepancias de nuestro esquema. La consecuencia es que la liberación de la dopamina desde sus terminaciones debe incrementar la fuerza de las sinapsis de algún modo. No se sabe cómo ocurre esto exactamente: puede ser que la liberación de la dopamina actúe directamente sobre los receptores sinápticos o puede actuar más indirectamente. En cualquier caso, lo que importa es que la activación de estas neuronas conduce de algún modo a una modificación de la fuerza de la sinapsis.
¿Le resulta beneficioso este sistema al mono? Para que el aprendizaje sea útil, tiene que estar relacionado con los actos del mono. Esto se puede conseguir al conectar el impulso saliente de la neurona de discrepancias y los movimientos del brazo del mono, lo que se muestra con la flecha de la figura 55. Inicialmente, los movimientos del brazo del mono no están relacionados con el sonido de la apertura de la puerta, pero después del condicionamiento, el mono mueve su brazo cuando lo escucha. Según el esquema de aprendizaje DT, esto ocurre porque al aumentar la activación de la neurona de discrepancias, se estimulan las acciones que probablemente incrementen la probabilidad de recoger una recompensa. Se trata de un proceso que se puede basar en un refuerzo sináptico parecido al descrito para la neurona de expectativas, con la excepción de que ahora las conexiones neurales serán las responsables de determinadas acciones, en vez de que las inicien las señales externas (en el próximo capítulo retomaremos la manera de reforzar las acciones). El resultado neto es que la estimulación de la neurona de discrepancias mediante el sonido de la puerta estimula el movimiento del brazo del mono para recoger la comida.
Antes de terminar esta explicación del aprendizaje DT, debo aclarar algo que he explicado por encima. He supuesto que la activación de la neurona de expectativas cae al mismo tiempo que se entrega la recompensa. Esto asegura que el efecto inhibidor de la neurona de expectativas contrarresta el efecto estimulador de la recompensa. Pero ¿por qué la activación de la neurona de expectativas debe caer exactamente cuando se entrega la recompensa? Según los partidarios del aprendizaje DT, la razón es que el cerebro posee la clave para discernir entre el momento o la duración de las señales, no solo su intensidad. Por ejemplo, podríamos imaginar que el sonido de la puerta pone en marcha en el cerebro una serie de patrones de activación con distinta duración: algunos duran poco tiempo y otros mucho[65]. Sería como si el sonido de la puerta reverberara durante un tiempo en el cerebro y proporcionara un abanico de impulsos entrantes para la neurona de expectativas. Supongamos que pudiéramos conseguir que solo se refuerce la sinapsis de la señal de entrada que dura hasta justo antes de que se entregue la recompensa, por lo que las señales de entrada más cortas o más largas no se verían fortalecidas por la neurona de discrepancias. Este sistema aseguraría que la activación de la neurona de expectativas caerá al mismo tiempo que llega la señal de recompensa, porque solo las señales con esta propiedad se verán fortalecidas.
Los mecanismos temporizadores de este tipo pueden sonar a elucubraciones, pero explican una observación neural clave. Supongamos que después del condicionamiento, al mono se le retira la recompensa. El animal meterá la mano en la caja, pero le disgustará que no haya una manzana. Schultz y colaboradores encontraron que, en esta situación, la tasa de activación de las neuronas dopaminérgicas cae exactamente en el momento en el que el mono suele esperar el contacto con la manzana. En otras palabras, la activación se inhibía justo cuando el mono esperaba recibir la recompensa. Este es el resultado que cabría esperar según el esquema de aprendizaje DT, porque la activación de la neurona de expectativas cae exactamente en este momento y provoca la inhibición de la neurona de discrepancias que ahora no estará compensada por una señal de recompensa (no hay manzana), por lo que la tasa de activación de la neurona de discrepancias cae por debajo de su valor normal. La reducción de la activación de la neurona de discrepancias se corresponde con la decepción del mono, al igual que el incremento de la activación se corresponde con una sorpresa agradable.
§. Pávlov y los castigos
El aprendizaje DT proporciona un mecanismo general para el aprendizaje predictivo, en el que interviene la interacción neural entre las expectativas, las recompensas y las discrepancias. El mismo esquema puede explicar el comportamiento del perro de Pávlov: al principio, la comida aparece como caída del cielo, sin que el perro se lo espere. Esta discrepancia entre la expectativa y la recompensa fortalece las conexiones sinápticas de las señales que la preceden, como el sonido de la campana. Gracias a este proceso aumenta la activación de las neuronas de expectativas del perro cuando escucha la campana, lo que conduce a un desplazamiento de la actividad de la neurona de discrepancias desde el momento de entrega de la comida al momento en que suena la campana. La principal diferencia respecto al ejemplo del mono es que esta neurona de discrepancias estimula el incremento de la salivación en vez de los movimientos del brazo. En realidad, el perro de Pávlov es más simple que el caso del mono porque no se tiene que aprender la conexión entre la discrepancia y la acción porque la respuesta de salivación ya estaba instaurada antes del condicionamiento. Los casos como el perro de Pávlov en los que la acción ya existía se denominan condicionamiento clásico. Por el contrario, los casos como el del mono que recoge la manzana, en los que la acción también se tiene que aprender, se denominan condicionamiento operante o instrumental[66]. El aprendizaje DT consigue explicar ambas clases de condicionamiento.El aprendizaje DT también es capaz de explicar por qué el condicionamiento con una señal predictiva, como tocar la pata, bloqueará el condicionamiento posterior con otra señal, como el sonido de la campana. Después de haber aprendido una señal predictiva, como tocar la pata, ya no se producirán más discrepancias en el momento de la entrega de la recompensa (la neurona de discrepancias ya no se estimula en ese momento). Por lo tanto, no habrá ningún refuerzo de las sinapsis con las señales que se produzcan por el toque de la pata. El sistema ya ha aprendido lo que ha de esperar y no fortalece las conexiones con los impulsos que no tienen ningún otro valor predictivo.
Hasta ahora he descrito ejemplos con recompensas como la comida. Pero el mismo esquema se puede aplicar a los castigos, lo opuesto a los premios. Supongamos que después de un destello, el mono recibe una descarga eléctrica en vez de un trozo de manzana si mete la mano en la caja: el mono aprende pronto a no meter la mano en la caja después de un destello de luz. Podemos aplicar el mismo esquema de aprendizaje DT que antes para conseguir este resultado, pero con un tipo diferente de neurona de discrepancias. En vez de recibir impulsos de recompensa, como una manzana, esta neurona de discrepancias recibiría impulsos repelentes como la descarga eléctrica. Otra diferencia es que la acción resultante de la activación de la neurona de discrepancias sería la evasión, no el compromiso. El aprendizaje proseguiría como antes, salvo que se inhibirían las acciones que ocurren inmediatamente antes de la activación de la neurona de discrepancias. Por lo tanto, si hay un destello, el mono aprendería a no meter la mano en la caja. Aunque en principio este esquema debería funcionar, las neuronas que intervienen todavía no se han identificado con claridad. Por lo tanto, conocemos menos detalles neurales del castigo que del aprendizaje con recompensas[67].
El aprendizaje DT proporciona hoy en día el mejor modelo para explicar las bases del aprendizaje predictivo mediante las interacciones neurales. Ahora quiero utilizar este ejemplo para fijarme en los principios básicos que intervienen en el aprendizaje. A partir de ahora utilizaré términos que también empleé al describir la evolución y el desarrollo para resaltar los principios fundamentales comunes, al mismo tiempo que reconoceré que estos procesos difieren también en muchos aspectos.
§. Principios fundamentales
Los cambios principales en la fuerza de la sinapsis no ocurren al mismo tiempo durante el aprendizaje DT, sino que dependen del efecto de la acumulación de muchas experiencias. Estamos tratando con una población de sucesos, con muchos casos de apertura de puertas y entrega de premios. Tal y como ya hemos visto, una población siempre se encontrará en un contexto, al igual que los giros de la ruleta o las moléculas en una taza de té. En el caso del aprendizaje DT, el contexto consiste en el conjunto de conexiones neurales del cerebro y el abanico de experiencias vividas. Cada experiencia por sí sola ocasionaría un único y ligero cambio de la fuerza sináptica, pero un conjunto de experiencias ocasionará un cambio significativo. Si no fuera así (si un único suceso fijara la fuerza sináptica), un animal formaría sus conexiones neurales solo de acuerdo con lo último que hubiera vivido, se re cablearía según los sucesos más recientes y borraría cualquier experiencia anterior. Pero la experiencia más reciente no nos da necesariamente el mejor pronóstico de lo que ocurrirá después, igual que ganar una vez a la ruleta no significa que ganaremos la siguiente. Hay algunos casos, en particular cuando recibimos un castigo fuerte, en los que una única experiencia puede bastar para que aprendamos, lo que se expresa con refranes como «gato escaldado del agua fría huye» o «la letra con sangre entra». En nuestro mundo complejo e impredecible suele ser mejor aprender sobre la base de tendencias globales y no tan solo con el pasado más reciente. Cuanto más inseguros estemos de nuestro entorno, bien por la falta de experiencia o por la variabilidad del mismo, más cuesta aprender de esta manera. Esta forma de aprendizaje es precisamente lo que consigue el aprendizaje DT, que ajusta ligeramente la fuerza de las sinapsis con cada acontecimiento, lo que permite aprender de tendencias predictivas en vez de un simple acontecimiento. El aprendizaje DT se construye sobre el principio de la variabilidad de la población.El aprendizaje DT también depende de la persistencia. El sistema no funcionaría si tras cada modificación de la fuerza de una sinapsis se volviera al estado inicial. Como vimos en el capítulo anterior, los cambios de la fuerza sináptica pueden persistir durante mucho o poco tiempo. La primera secuencia de experiencias durante el aprendizaje podría provocar cambios a corto plazo en el funcionamiento de la sinapsis, haciéndola más o menos eficaz. Estos cambios a corto plazo se acumularían con otras experiencias para dar lugar a cambios anatómicos a más largo plazo, como el número de sinapsis[68]. Los cambios sinápticos a más largo plazo consiguen durar muchos años, como se ilustra en la siguiente anécdota de Charles Darwin.
En 1836, justo después de regresar de sus cinco años de travesía a bordo del Beagle, Darwin decidió comprobar la memoria de su perro[69]. Se fue al establo en el que estaba alojado el perro y lo llamó. El perro se levantó corriendo y se apresuró, feliz, para dar un paseo con Darwin, con la misma emoción que si su amo hubiese faltado tan solo media hora. El sonido de la voz de Darwin seguía estimulando la respuesta del perro después de cinco años de ausencia. Quizá esta historia también nos dice algo del cerebro de Darwin: había aprendido literatura clásica en el colegio, donde solía memorizar cuarenta o cincuenta líneas de Homero antes de rezar por la mañana en la capilla[70]. En la historia clásica de Homero, La Odisea, Ulises regresa a su hogar después de diez años de viaje marítimo. Allí se disfraza de mendigo para impedir que lo reconozcan, pero no consigue engañar a su viejo perro Argos, que instantáneamente levanta las orejas y empieza a menear la cola en cuanto suena la voz de su amo. Darwin pensaba que recitar Homero en el colegio era una total pérdida de tiempo porque olvidaba cada verso al cabo de dos días. Pero quizá el relato de Ulises estableció algunas conexiones neurales a largo plazo en el cerebro de Darwin que le estimularon inconscientemente nada más regresar de su largo viaje marítimo, y eso quizá le condujo a comprobar la memoria de su perro. Darwin y su perro quizá se parecían más incluso de lo que él creía.
La variabilidad de la población y la persistencia son ingredientes clave del aprendizaje: sin variación no habría cambios en la fuerza de las sinapsis, y sin persistencia, cada cambio sináptico se desvanecería en cuanto hubiera acabado de formarse.
El refuerzo también desempeña una función vital en el aprendizaje DT. La activación de la neurona del sonido de la puerta fomenta la fuerza de su propia sinapsis, con la ayuda de la neurona de discrepancias. Tal refuerzo resulta esencial para que funcione el sistema, pues gracias a él, a la hora de estimular la respuesta del mono, la neurona del sonido de la puerta se vuelve más eficaz que otras posibles señales como un destello. De igual forma, con el perro de Pávlov, la activación de una neurona para el sonido de la campana tiende a reforzar sus propias sinapsis. Podemos resumir tal refuerzo con un bucle de realimentación positiva tal y como se muestra a la izquierda de la figura 56, donde la activación de una neurona favorece su propio refuerzo sináptico.
Si se las tuviera que arreglar solo, el refuerzo haría que determinadas sinapsis fueran cada vez más fuertes a medida que se acumularan las experiencias. El sistema intensificaría por sí mismo cada vez más la actividad sináptica, lo que llevaría al cerebro finalmente a un estado de frenesí. Esto no ocurre porque el auto refuerzo acaba siendo víctima de su éxito: a medida que la sinapsis con la neurona del sonido de la puerta incrementa su fuerza, la activación de la neurona de expectativas también aumenta al escuchar el sonido de la puerta. En consecuencia, la neurona de expectativas se activa mucho menos cuando se entrega la recompensa, lo que va extinguiendo la actividad de la neurona discrepante y, por lo tanto, cualquier otro refuerzo se ve reducido. El refuerzo ha conducido a su propia inhibición o limitación, como se muestra mediante el bucle negativo a la derecha de la figura 56. Se trata de nuestro conocido doble bucle de realimentación en el que el refuerzo promueve la competencia, y en el que la competencia afecta a la acción inhibidora de las neuronas. El efecto global de esta combinación de bucles negativos y positivos es que las expectativas se pueden hacer coincidir con los niveles de recompensa o castigo.
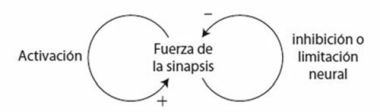
Figura 56. Interacción entre el refuerzo (bucle positivo) y la competencia (bucle negativo) en el aprendizaje.
Por supuesto, hay muchas diferencias entre la evolución, el desarrollo y el aprendizaje. Las diferentes experiencias no son lo mismo que las variaciones genéticas en una población ni las colisiones entre moléculas, y el refuerzo de las sinapsis es muy diferente del éxito reproductor. A pesar de las cuantiosas diferencias, hay no obstante algunas similitudes de forma a un nivel superior, a saber, un conjunto parecido de bucles de realimentación e ingredientes básicos.
No es una casualidad que hayamos llegado a algunos principios comunes. Después de todo, la estrategia que estamos siguiendo en este libro ha consistido en fijarnos en lo que podrían compartir las diferentes transformaciones. Así pues, el objetivo de este ejercicio no ha sido buscar las similitudes porque nos gusten, sino para ayudarnos a comprender la esencia de las transformaciones de los seres vivos. Las explicaciones tradicionales del aprendizaje DT no lo presentan como un doble bucle de realimentación entre el refuerzo y la competencia, impulsado por un equilibrio de variabilidad y persistencia. Pero viéndolo desde esta perspectiva, apreciamos con más claridad tanto la lógica fundamental del aprendizaje DT como su relación con otros procesos.
He presentado el aprendizaje DT como ejemplo para ilustrar que los principios comunes también valen para un determinado mecanismo de aprendizaje. Pero se podría decir lo mismo de la mayoría de los mecanismos que se han propuesto para el aprendizaje[71]. En un entorno variable e impreciso necesitamos aprender las tendencias de una población de sucesos y no de ejemplos únicos. Para aprender a obtener efectos duraderos, tiene que haber cierta forma de persistencia. Para que algunas conexiones neurales se amplifiquen a sí mismas más que a otras, tiene que haber refuerzo. Y el refuerzo debe auto limitarse mediante la competencia para hacer coincidir nuestras experiencias y lo que se aprende. Los detalles pueden variar de un mecanismo de aprendizaje a otro, pero siempre estarán ahí los mismos principios fundamentales.
§. Un periplo neural
Para aprender se necesita más de una neurona. En el caso del aprendizaje DT existen interacciones entre las neuronas de expectativas y de discrepancias, así como entre las neuronas que perciben estímulos como el contacto, la luz y el sonido, y las neuronas que intervienen en las acciones como los movimientos del brazo o la salivación. Estas neuronas también pueden ayudar a las acciones de las otras. El refuerzo de la sinapsis entre la neurona del sonido de la puerta y la neurona de expectativas depende no solo de la activación de la neurona del sonido de la puerta, sino también del ritmo al que se activa la neurona de discrepancias. Para el refuerzo sináptico se necesita la concomitancia de ambas activaciones, de manera que los impulsos que salgan de las neuronas del sonido de la puerta y de discrepancias deben converger sobre la neurona de expectativas. Tal y como hemos visto antes, la cooperación también depende de la proximidad física, pues habrán de reunirse determinadas terminaciones neurales y dendritas. Sin esta clase de proximidad, cada neurona funcionaría de forma independiente y sería imposible aprender nada. El principio de cooperación es otro ingrediente clave del aprendizaje.La reunión de varios componentes abre las puertas a numerosas combinaciones. El cerebro humano contiene aproximadamente 100.000 millones de neuronas, cada una de las cuales puede tener miles de sinapsis, por lo que el número total de conexiones sinápticas es unas mil veces mayor: cien billones. Esto abre un enorme espacio de posibilidades neurales. Ya nos hemos encontrado grandes espacios: al describir la evolución nos topamos con el espacio genético, o sea, el abanico de secuencias de ADN posibles; y para el desarrollo nos encontramos con el espacio del desarrollo, el abanico de estados celulares posibles y su organización en un embrión. Todos ellos eran hiperespacios inmensos con numerosas dimensiones. De igual forma, ahora tenemos espacio neural, el enorme conjunto de conexiones posibles y de estados de activación de las neuronas del cerebro. Se trata del principio de la riqueza combinatoria aplicado a las conexiones neurales.
Anteriormente hemos pensado en la evolución como poblaciones que viajaban por el espacio genético, y en el desarrollo como un embrión que viajaba por el espacio del desarrollo. De igual forma podríamos pensar que el aprendizaje es un cerebro que viaja por el espacio neural. Cada uno hemos nacido con un cerebro muy estructurado, que corresponde a una posición particular de este espacio neural imaginario. Esta posición es ligeramente diferente para cada uno de nosotros, en buena parte debido a las diferencias en la composición genética, que influyen en el desarrollo del cerebro desde el útero. Después del parto, comenzamos a interaccionar con fuerza con nuestro entorno, de manera que cada interacción desplaza nuestra posición en el espacio neural y nos lleva a emprender un periplo neural muy intrincado. El aprendizaje es en buena parte responsable de nuestro periplo de desplazamientos al cambiar la fuerza y el número de las conexiones neurales. Los ejemplos del perro de Pávlov y del mono que recoge su recompensa ilustran la manera en que las conexiones consiguen cambiar gracias a unas pocas interacciones entre las neuronas. Pero la misma clase de cambios ocurrirán en paralelo en numerosas entradas, salidas y conexiones del cerebro, lo que nos guiará por nuestro elaborado periplo neural.
Una de las principales consecuencias de este periplo neural es que mejora nuestra capacidad para predecir lo que es probable que suceda. Continuamente modificamos nuestras expectativas sobre la base de la historia de nuestras experiencias, gracias a lo cual somos capaces de afrontar mejor nuestro entorno. Pero ¿qué nos mantiene en movimiento por el espacio neural? ¿Qué nos proporciona la fuerza impulsora continua para el aprendizaje?
§. Quedarse en marcha
Una de las características clave del aprendizaje DT es que no elimina las discrepancias, simplemente las desplaza. Para el experimento del mono, el resultado global es que el sonido de la puerta, y no el tocar la manzana, viene a estimular la neurona de discrepancias, por lo que la discrepancia se desplaza a un momento anterior. Mientras que, antes, el mono se excitaba al tocar la manzana, ahora se excita al escuchar el sonido de la puerta. Según los defensores del aprendizaje DT, esto no es más que una consecuencia de que la activación de la neurona de discrepancias excita o da placer al mono. De hecho, el neurotransmisor dopamina liberado por la neurona de discrepancias (como estudiaron Romo y Schultz) se piensa que desempeña una función importante en la toxicomanía. Las drogas como la cocaína y las anfetaminas se cree que ejercen su efecto al potenciar la acción de la dopamina. Inicialmente, tocar la manzana estimulaba la neurona de discrepancias y la liberación de dopamina, con lo que se excitaba el mono. Después del aprendizaje, el sonido de la puerta se comporta igual que la manzana al principio: estimula la neurona de discrepancias y la liberación de dopamina. Desde esta perspectiva neural, el sonido de la puerta se ha convertido en un sustituto de la recompensa, al estimular la neurona de discrepancias y la liberación de dopamina igual que hacía la manzana. Si se produce un nuevo estímulo, como un destello luminoso, antes del sonido de la puerta, entonces el mono trataría esta situación como si el destello sirviera de pronóstico de la recompensa. Pero en este caso, la «recompensa» no es el contacto con la manzana, sino su sustituto: el sonido de la puerta. Si el mono experimenta muchas veces el destello antes del sonido de la puerta, el aprendizaje DT garantiza que la neurona de discrepancias empieza a activarse incluso antes, en el momento del destello. Estamos construyendo una expectativa sobre otra. Al haber aprendido a esperar la recompensa de la manzana sobre la base del sonido de la puerta, el sistema neural automáticamente aprende a responder a factores que le permitirían predecir el sonido de la puerta. El impulso para aprender no se detiene, sino que ha cambiado a otro estímulo.La capacidad de aprender para crear así sustitutos de recompensas está tan extendida que puede ser difícil identificar lo que constituye una recompensa instintiva frente a una aprendida. En muchos experimentos de Pávlov, el olor o la visión de la comida se utilizaron para condicionar otras respuestas. Podríamos pensar que los perros tienen una respuesta de salivación instintiva ante la presencia de la carne y aprenden las señales que actúan como pronóstico de esta recompensa, como el sonido de la campana. Pero la respuesta a la carne resulta que no es instintiva, sino el resultado del condicionamiento[72]. Si un perro recién nacido se alimenta durante bastante tiempo únicamente con leche, entonces no saliva la primera vez que huele o ve la carne, un hecho que solo comenzará a evocar la reacción de salivación después de que el perro se haya alimentado con carne unas cuantas veces. Ahora el olor y la visión de la carne predicen la recompensa igual que una comida satisfactoria, y comienzan a convertirse en recompensas por su propio derecho. Lo que normalmente pensamos que es una reacción instintiva o no aprendida, como la salivación en respuesta a la carne, realmente surge a través del condicionamiento, pero esto no significa que el perro nazca sin ninguna respuesta instintiva. Los modos innatos de responder a los estímulos que actúan como premios o castigos están determinados por las conexiones neurales que se establecen durante el desarrollo del embrión de perro. Pero en cuanto nace y queda expuesto al entorno, comienza a construir sobre estas reacciones a través del aprendizaje, y establece otros conjuntos de estímulos que actúan como premios o castigos.
El fenómeno de que una respuesta aprendida constituya el fundamento para otra se denomina condicionamiento secundario, y surge de forma natural por el aprendizaje DT porque este mecanismo funciona de manera relacional. A medida que aparecen discrepancias, las expectativas cambian y desplazan a las discrepancias con la introducción de sustitutos de las recompensas para la siguiente ronda de aprendizaje.
Los humanos, con sus cerebros elaborados, son expertos en este juego de ajuste de expectativas y recompensas. A menudo consideramos que el dinero es una recompensa, por lo que trabajamos para que nos paguen a final de mes. Pero el dinero solo tiene valor porque nos permite comprar cosas como la comida u otros bienes. Podemos, por ejemplo, comprar una manzana en una tienda, y seguramente no nos la comeremos inmediatamente, sino que retrasamos aún más la recompensa consumiéndola más tarde. Cada una de estas recompensas (dinero, comprar comida y comer) está conectada a determinadas expectativas, de las que somos conscientes gracias a las discrepancias. Si nos pagan menos de lo que esperamos, notamos la discrepancia y quizá nos quejemos al empresario. O si entramos en una tienda esperando comprar una manzana, pero encontramos que su precio ha subido de repente, notamos el encarecimiento. Y si llegamos a casa y encontramos las manzanas más agrias de lo esperado, notamos otra discrepancia. Si todo va viento en popa y se satisfacen nuestras expectativas, entonces apenas reflexionamos sobre estos acontecimientos. El mundo se nos manifiesta gracias a las discrepancias[73].
Estas discrepancias están relacionadas con la predicción de consecuencias globales en el futuro. Probablemente nos sentiremos más decepcionados si nos recortan el salario que si sube el precio de las manzanas: una pérdida de ingresos probablemente nos resulte más adversa que el encarecimiento de unas pocas manzanas. Las discrepancias se miden frente a la suma de recompensas futuras: un cambio de salario se pondera frente a todos los bienes que podríamos comprar con esa cantidad de dinero, no con una determinada compra. Esta capacidad de aprendizaje respecto a la suma de las recompensas futuras constituye también un resultado natural de aprendizaje DT y es reflejo del modo relacional con el que trabaja. Las discrepancias en cualquier momento se relacionan con los cambios de expectativas y recompensas que las seguirán, lo que permite que la suma de los sucesos futuros influya en el aprendizaje.
El aprendizaje implica responder continuamente a las discrepancias y reajustar las expectativas. Si nos suben el sueldo, inicialmente nos sentimos positivos porque nuestra recompensa excede a las expectativas. Pero pronto aprendemos a esperar este nuevo salario y nos sentiríamos decepcionados si volviera a su nivel anterior. Lo mismo ocurre con los cambios de precio, o con la calidad de las manzanas que compramos. Se trata de sucesos quizá más sofisticados que la salivación del perro de Pávlov, o que el dinero alcance para comprar comida, pero los fundamentos neurales son los mismos: aprendemos de las discrepancias y reajustamos nuestras expectativas en consonancia, con lo que desplazamos las discrepancias durante dicho proceso.
Lo que mantiene nuestro periplo neural en constante movimiento es este aspecto relacional del aprendizaje que induce constantemente el desplazamiento de las discrepancias. Si pudiéramos eliminar de algún modo todas las discrepancias, entonces el aprendizaje se detendría completamente. Supongamos que pudiéramos ver perfectamente en el futuro y anticiparnos exactamente a lo que va a ocurrir. Ya no tendríamos ninguna sorpresa ni discrepancia de la que preocuparnos, con lo que no habría nada que aprender. Esto podría sonar a encontrarse en un estado ideal. Sin embargo, dado el modo en el que trabaja nuestro cerebro, nos entumecería la mente: sin nada que nos sorprenda, no habría nada que atrajera nuestra atención, ninguna discrepancia para ejercitar nuestra mente. Un conocimiento perfecto del futuro actuaría como un anestésico mental.
En la práctica, el riesgo de alcanzar tal omnisciencia es escaso porque, a pesar de que nuestro cerebro sea complejo y rico, el mundo que nos rodea es todavía más rico. Solo podemos aspirar a capturar una mínima fracción de la complejidad de nuestro entorno, por lo que siempre habrá discrepancias y problemas con los que tratar, y por eso encuentran trabajo las sibilas, los profetas y las pitonisas. El aprendizaje nunca eliminará por completo los desafíos, solo introduce otros nuevos. Las discrepancias nos resultan tan importantes que incluso las creamos para entretenernos. Las películas o los libros suelen terminar cuando todo el mundo es feliz, porque como no quedan discrepancias, ya no hay nada que nos enganche. Es una lástima, porque después de haber experimentado un episodio dramático, podríamos querer saborear la felicidad del final. Pero no queda nada que siga reteniendo nuestra atención una vez que todas las discrepancias de la narración se han resuelto, por lo que los finales felices tienden a ser mucho más cortos que la historia que los precede. Tólstoi nos planteó lo mismo al comienzo de su libro Anna Karenina: «Todas las familias felices se asemejan; cada familia infeliz es infeliz a su modo». La discordia atrae nuestro interés.
Lo mismo ocurre con la enseñanza: para que un alumno aprenda una nueva idea, debe haber una discrepancia a la que aferrarse. Si un alumno siente que ya lo sabe o no ve que haya ningún problema al que enfrentarse, entonces aprenderá poco. Un buen profesor tiene que ser capaz de estimular las discrepancias del alumno planteándole un problema o pregunta que le excite o que le enganche. Una vez que haya aprendido a resolver dicha discrepancia, la siguiente etapa de aprendizaje consistirá en introducir una nueva discrepancia, otro problema que resolver.
§. Una receta para el aprendizaje
Cada uno de nosotros va siguiendo un periplo neural concreto desde el mismo momento del nacimiento. El cerebro de un recién nacido ya lleva una serie de expectativas implantadas en sus conexiones e interacciones neurales. También tiene un conjunto de valores neurales, modos de responder a los estímulos que actúan como premios o castigos, que son el reflejo de su posición de partida en el espacio neural, posición que depende del modo intrincado en el que creció y se desarrolló el tejido neural del embrión, y del modo en el que se formó el cerebro con su compleja estructura. A medida que los recién nacidos comienzan a experimentar el mundo, se topan con las discrepancias y estas conducen a un desplazamiento de las expectativas, que los arrastran a nuevas regiones del espacio neural. Esto a su vez cambia el contexto neural e introduce nuevas discrepancias y nuevos valores conectados con los anteriores. Por lo tanto, el proceso continúa gracias a que el propio periplo neural resulta auto estimulador. El periplo neural para cada persona es único como resultado de que cada una empezó en una posición diferente del espacio neural y de que nuestras propias experiencias también difieren. No obstante, el periplo está impulsado por el mismo conjunto de procesos repetitivos que en el principio de la recurrencia, pero ahora sobre el dominio del aprendizaje. En la evolución, la recurrencia surge porque la selección natural funciona de manera relacional. A medida que las adaptaciones se diseminan y las comparte la población, todos los individuos son capaces de funcionar mejor, y esto conduce a la competencia por adaptaciones cada vez más eficaces. En el desarrollo, la recurrencia implica que los patrones de la actividad génica de un embrión en crecimiento vayan cambiando continuamente. A medida que se modifica un patrón, desplaza el contexto molecular y celular, lo que conduce a otro conjunto de activaciones génicas y transformaciones. En el aprendizaje, la recurrencia surge de otra manera diferente: a medida que se resuelven algunas discrepancias neurales, se establecen otras que entonces conducen a nuevas rondas de aprendizaje. En todos los casos, el principio de la recurrencia es lo que mantiene el periplo en marcha.Al igual que la evolución y el desarrollo, el aprendizaje se fundamenta en un conjunto de principios comunes que interaccionan entre sí, aunque ahora se hayan disfrazado de neuronas. El núcleo del aprendizaje implica un bucle doble de realimentación en el que la activación neural se refuerza a sí misma, además de acarrear sus propios límites debido a la competencia. Estos bucles están siendo alimentados por la variación de la tasa de activación provocada por un conjunto de experiencias, lo que conduce a la persistencia de los cambios de fuerza y del número de las conexiones sinápticas. Esto sucede gracias a las interacciones cooperativas entre muchas neuronas que crean colectivamente un enorme espacio de combinaciones posibles. Cada etapa del aprendizaje conduce a un desplazamiento por el espacio neural, con la construcción recurrente sobre el contexto neural ya existente, con lo que se modifica el cerebro y se desplaza el contexto. El mismo conjunto de principios e interacciones interviene en el núcleo del aprendizaje, como en la evolución y en el desarrollo. El aprendizaje es nuestra tercera manifestación de la receta creativa para la vida. Pero en vez de impulsar los organismos por el espacio genético o del desarrollo, en este caso, la receta acompaña a los organismos por un periplo a través del espacio neural.
Capítulo 8
Aprendizaje mediante acciones
§. CalibraciónHemos visto que al igual que la evolución y el desarrollo, el aprendizaje es una manifestación de la receta creativa para la vida. Mi explicación del aprendizaje podría detenerse en este punto ya que hemos estudiado los principios fundamentales que intervienen. Sin embargo, si acabo aquí, dejaría muchas preguntas sin respuesta: ¿cómo están conectados los principios que hemos tratado con procesos como la memoria, el movimiento, la atención, el reconocimiento, el lenguaje, la creatividad y la consciencia? Cada uno de estos aspectos del conocimiento ha sido objeto de intensa investigación, pero por desgracia todavía no sabemos muchos detalles neurales sobre la manera en que el aprendizaje se incorpora en ellos. Incluso así, los principios que nos hemos encontrado proporcionan un marco de trabajo o unas herramientas conceptuales para comprender el funcionamiento del aprendizaje en tales casos. Si vemos la cognición de este modo, podremos explorar algunas otras ramificaciones del aprendizaje, y también fijar el escenario para centrarnos en los cambios culturales en los capítulos siguientes.
§. Ojos saltarines
§. Desplazamientos visuales
§. Aprender a calibrar
§. Bucles de aprendizaje y acción
§. Movimientos suaves
§. Un periplo activo
§. Se aprende con otros
No me sería posible abarcar todas las áreas del conocimiento, por lo que he preferido concentrarme en unos pocos aspectos clave para ilustrar algunos de sus principios fundamentales. En este capítulo veremos que el aprendizaje y las acciones están entrelazados, mientras que en el capítulo siguiente exploramos la manera en que aprendemos a ver las cosas de un modo particular. En esta descripción utilizaré con frecuencia ejemplos de la vista, pero los principios generales también valdrán para otras modalidades sensoriales. Al estudiar el conocimiento a través de estos principios, veremos que nuestros periplos neurales no se mueven por un impulso pasivo, sino que están conectados íntimamente con nuestras acciones e interpretaciones.
Dada nuestra falta de conocimiento directo en algunas áreas, utilizo algunos esquemas hipotéticos para ilustrar los posibles mecanismos subyacentes. Aunque esta estrategia introduzca un cierto nivel de especulación, tiene el mérito de que, al perseguir ideas concretas, podremos apreciar mejor dichos principios subyacentes. Sin embargo, algunos lectores quizá prefieran saltarse las explicaciones más detalladas en estos dos capítulos. Tratemos primero la calibración, uno de los modos más básicos con que aprenderemos a conectar las acciones con sus efectos.
§. Calibración
Leonardo da Vinci estaba familiarizado con los conflictos militares. Durante su vida, Italia estuvo bajo la continua amenaza de invasión desde Francia, y Florencia, su ciudad natal, entraba en guerra con su vecina, Pisa, con cierta regularidad. Por lo tanto, no es sorprendente que Leonardo a menudo pusiera su mente inventiva a trabajar en máquinas de guerra. Al igual que concibió invenciones militares, como el cañón multibarril, Leonardo se interesó por la ciencia de los proyectiles, o balística. Se hacía preguntas sobre la potencia de lanzamiento de una bombarda (un tipo de cañón): «Si con cuatro libras de pólvora una bombarda lanza a dos millas una bola de cuatro libras, ¿adónde llegará si se lanza con seis libras de pólvora?»[74]. La respuesta a esta cuestión no solo tenía un interés teórico, sino también práctico: si queremos acertar en una diana militar mediante una bombarda, necesitamos saber cuánta pólvora utilizar.El problema balístico planteado por Leonardo se puede abordar de varias maneras. Una estrategia práctica consistiría en anotar la distancia que alcanzó la bola lanzada por una bombarda alimentada con distintas cantidades de pólvora. Esto nos daría una tabla de calibración, una lista de distancias para cada cantidad de polvo que se utilizara. Entonces podríamos determinar cuánta pólvora se necesitaría para la distancia requerida con tan solo buscar en la tabla. Una limitación de esta estrategia consiste en que solo podríamos buscar distancias que ya se hubieran comprobado en los ensayos. Si nuestra distancia no estuviera recogida en la tabla, no sabríamos cómo seguir. Una mejora sería construir una curva de calibración, un gráfico de la distancia recorrida frente a la pólvora utilizada. Entonces podríamos buscar cuánta pólvora necesitamos para alcanzar cualquier distancia simplemente examinando dónde corta la curva.
Figura 57. Bombardas dibujadas por Leonardo da Vinci, hacia 1504.
§. Ojos saltarines
La acción más frecuente que realizamos en la vida consiste en mover los ojos: cuando miramos una escena, los ojos saltan continuamente de un lugar a otro. En la figura 58 (lámina 9) se muestran estos movimientos balísticos de los ojos, o sacudidas oculares, de alguien que contempla Bodegón con tetera de Cézanne. En muy poco tiempo, el ojo salta por el cuadro muchas veces, y se posa con mucha frecuencia cerca de los objetos, como la manzana o el borde de la vasija. Estos movimientos oculares son más frecuentes que los latidos del corazón porque realizamos unas tres sacudidas oculares cada segundo, lo que equivale a varios miles de millones durante la vida. Algunas corresponden a saltos relativamente grandes, como el desplazamiento desde la manzana a la vasija. En cambio, otras no son más que pequeñas agitaciones alrededor de un punto (se denominan micro sacudidas oculares). Apenas somos conscientes de la mayor parte de estas sacudidas, pues se producen inconscientemente, dirigidas por la actividad del cerebro. Las sacudidas oculares no son solo una particularidad de la vista humana, sino que también las presentan muchos otros animales, desde los calamares a la carpa dorada[75].Las sacudidas oculares tienen su origen principalmente en una limitación de los ojos relacionada con la existencia de muchos más fotorreceptores concentrados en la región central de la retina, conocida como la fóvea, en donde la vista es mucho más aguda que en la periferia. Con el movimiento continuo del ojo y el desplazamiento de la mirada, obtendremos una gran definición de las diferentes áreas de una escena ante nosotros, lo que nos proporciona más información sobre la escena. Una consecuencia es que la imagen sobre la retina está cambiando continuamente mientras miramos una escena fija, como un cuadro. En lo que respecta a la retina, la situación es la misma que si alguien estuviera continuamente zarandeando el cuadro mientras lo contemplamos con la mirada fija. Aun así, no percibimos la agitación del cuadro a medida que desplazamos nuestra mirada sobre él. ¿Por qué?

Figura 58. Movimientos oculares registrados durante veinte segundos mientras se observa Bodegón con tetera, de Paul Cézanne, hacia 1869. Véase la lámina 9.
La prueba de que calibramos los ojos mediante el aprendizaje procede del siguiente experimento[77]: si sentamos a alguien en una habitación oscura y le pedimos que siga un punto de luz a medida que salta de una posición a otra, sus ojos lo seguirán. Cada vez que se desplace la luz, los ojos detectarán el cambio y saldrán disparados de tal forma que se posarán para que la luz se encuentre en el centro del campo visual. Estos movimientos oculares se pueden vigilar electrónicamente, lo que permitirá realizar un truco visual. A medida que los ojos salen disparados para seguir el objetivo, este se hace retroceder un poco antes de que se posen. Al principio, como cabría esperar, los ojos no darán en el blanco, pero después de unos pocos ensayos se adaptan automáticamente y aprenden a dar saltos ligeramente más pequeños; entonces volverán a posarse sobre el objetivo correctamente, incluso aunque retroceda. Los ojos se recalibran por sí solos a la nueva situación.
Tiene sentido aprender a calibrar porque cada uno de nosotros ha nacido con cuerpos y fuerzas musculares ligeramente diferentes, que a su vez irán cambiando a medida que crecemos. Por ejemplo, la potencia de los músculos oculares del lector puede ser ligeramente diferente de la mía. Si nuestras calibraciones fueran instintivas y fijas, entonces sería difícil verlas funcionar con fiabilidad cuando se enfrentan con situaciones variables. Del mismo modo, si cada bombarda funcionara de manera un poco diferente debido a las variaciones de fabricación, tendría sentido calibrar cada una individualmente según su rendimiento.
§. Desplazamientos visuales
¿Qué clase de interacciones neurales podrían ser las responsables de la calibración visual? Comencemos con una escena estacionaria y veremos que nuestras propias acciones, los movimientos de los ojos, están relacionadas con los efectos visuales que generan. Imaginemos que estamos mirando un objeto gris sobre un fondo negro con la cabeza quieta. El panel superior de la figura 59 representa lo que ocurre en el fondo de un ojo. Para simplificarlo un poco, he representado todo en una dimensión. La llegada de la imagen del objeto a la retina está indicada por una línea gris, mientras que la retina está representada por debajo de esta como una sola fila de fotorreceptores, o sea, de neuronas que pueden estimularse por la luz. He colocado la imagen del objeto gris en el centro del campo visual. Podemos observar que solo se estimulan con fuerza los fotorreceptores que reciben la luz del objeto, lo que está indicado con las líneas verticales dentro de los fotorreceptores.El panel inferior muestra lo que sucede si desplazamos la retina a la derecha con un movimiento ocular impulsado por una neurona que se muestra en gris oscuro, y que también estimula la contracción de algunos músculos oculares. Supondremos que cuanto mayor sea la tasa o la duración de la activación de esta neurona de movimientos oculares, más se desplaza la retina a la derecha (por motivos de simplicidad, ignoro la inversión óptica que tiene lugar cuando la luz entra en el ojo). Cuando la retina se mueve a la derecha, cambia la actividad de sus fotorreceptores: algunos que estaban antes en la oscuridad ahora reciben luz del objeto gris, mientras que otros que estaban estimulados por la luz ahora están en la oscuridad. Nuestro primer paso consiste en intentar relacionar este cambio de los impulsos visuales con la magnitud del desplazamiento del ojo. Queremos encontrar neuronas visuales que respondan en proporción a la magnitud del movimiento ocular.
Fijémonos en un único fotorreceptor en la parte izquierda de la retina, señalado con una flecha blanca en la figura 59. Este fotorreceptor estaba mudo al principio, pero se estimuló cuando se desplazó la retina porque el objeto gris entró en su campo visual. ¿Es la actividad de este fotorreceptor proporcional al desplazamiento ocular? El problema reside en que este fotorreceptor se ve estimulado con la misma intensidad por desplazamientos más grandes que el mostrado. Aunque la alteración de la actividad de este fotorreceptor nos dice que hay cierta clase de cambio, la actividad no es proporcional al desplazamiento del ojo. Lo mismo ocurre con cualquier otro fotorreceptor, porque ningún fotorreceptor por sí solo responderá proporcionalmente al grado de movimiento del ojo.
Figura 59. Objeto (línea horizontal gris) observado por la retina antes (panel superior) o después (panel inferior) del desplazamiento hacia la derecha de la retina impulsado por una neurona de movimientos oculares. La longitud de las líneas verticales dentro de los fotorreceptores indica el nivel de la actividad neural.
Figura 60. Combinación de información visual procedente de los fotorreceptores. En el panel inferior, la retina se ha desplazado hacia la derecha por la activación de una neurona de movimientos oculares. La longitud de las líneas verticales dentro de los fotorreceptores indica la magnitud de la actividad neural.
Supongamos que nuestro cerebro es capaz de utilizar información como la proporcionada por las neuronas ABC para estimular lo que denomino una neurona de desplazamientos visuales, que se muestra en el panel inferior de la figura 60. Podemos disponer las cosas de tal modo que si no hubiera ningún cambio de los impulsos procedentes de la retina, no se active la neurona de desplazamientos visuales; y que cuanta más retina se desplaza a la derecha, más se activa la neurona de desplazamientos visuales. También suponemos que nuestra neurona de desplazamientos visuales se comporta de este modo independientemente del tipo de imagen presentada. He ofrecido un ejemplo en el que se contempla un objeto gris sobre un fondo negro, pero vamos a suponer que la neurona de desplazamientos visuales responde de forma parecida a muchas otras imágenes. Al mostrar una determinada imagen, nuestra neurona de desplazamientos visuales se activa según la cantidad de imagen que se ha movido posteriormente por la retina[78]. La manera de conseguir exactamente esto no nos debe preocupar aquí, puesto que lo realmente importante es que la información visual combinada procedente de neuronas como las ABC se utiliza de alguna forma para generar una señal que se correlaciona con el movimiento ocular (estrictamente hablando, la señal se responde con los movimientos angulares, porque la activación de la neurona de desplazamientos visuales depende del desplazamiento angular ocasionado por la rotación del ojo).
§. Aprender a calibrar
Ahora tenemos dos cambios correlacionados: la activación de la neurona de movimientos oculares y la activación de la neurona de desplazamientos visuales, con lo que conseguimos conectar los mundos de la acción y de la percepción. Cuantas más neuronas de movimientos oculares se activen, más se desplaza el ojo a la derecha (acción), lo que a su vez cambia la entrada visual y conduce a un incremento de la activación de la neurona de desplazamientos visuales (percepción). Ahora quiero mostrar cómo se podría calibrar este sistema. En vez de construir tablas y dibujar líneas como en el caso de la bombarda, la calibración se consigue mediante las interacciones entre las neuronas.Miremos la figura 61, en la que se muestra el mismo esquema que antes, pero con una neurona de discrepancias adicional. Recordemos del capítulo anterior que las neuronas de discrepancias están implicadas en la detección de discordias o desfases. Para apreciar que esto podría ayudarnos con los movimientos oculares, primero miremos a lo que la neurona de movimientos oculares está realizando en nuestro esquema. Podemos ver que hace dos cosas: envía una señal para impulsar el movimiento del ojo y envía la misma señal (conocida como la copia eferente) a la neurona de discrepancias. La señal hacia la neurona de discrepancias se envía con retraso para que llegue después de que haya ocurrido el movimiento del ojo, y entonces estimula a la neurona de discrepancias a través de una sinapsis estimulante fuerte (signo positivo).
La señal desde la neurona de desplazamientos visuales también va a la neurona de discrepancias, donde llega al mismo tiempo que la señal retrasada desde la neurona de movimientos oculares. Sin embargo, a diferencia de la neurona de movimientos oculares, la neurona de desplazamientos visuales está conectada a la neurona de discrepancias por una sinapsis inhibidora (signo negativo), por lo que al activarse, silencia a la neurona de discrepancias. Además, la fuerza de esta sinapsis con la neurona de desplazamientos visuales no está fijada, sino que puede cambiar según la realimentación desde la neurona de discrepancias (flecha gris hacia arriba). Si la neurona de discrepancias se activa por encima de su tasa basal, entonces tenderá a reforzar la sinapsis. Y a la inversa, si la neurona de discrepancias se activa por debajo de su tasa basal, debilitará la sinapsis. Tenemos un sistema de realimentación, donde la señal emitida por la neurona de discrepancias influye en la fuerza del impulso que recibe desde la sinapsis con la neurona de desplazamientos visuales.
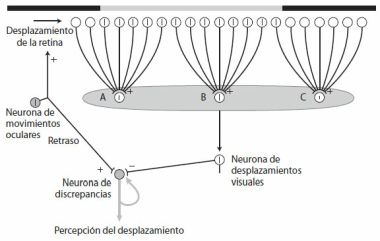
Figura 61. Aprendizaje de la forma de calibrar con una neurona de discrepancias.
Después de haber calibrado así el ojo, el sistema puede decirnos si un desplazamiento visual está provocado por el movimiento ocular o por un acontecimiento externo. Si el ojo se mueve a la derecha, la neurona de discrepancias se activa a su tasa basal debido a la calibración anterior. Pero supongamos que el mismo cambio visual de nuestra retina venga provocado por un acontecimiento externo que desplaza la escena, como cuando se mueve el cuadro. La neurona de discrepancias no recibe ningún impulso desde la neurona de movimientos oculares (los ojos no se han movido), aunque recibe un impulso inhibidor desde la neurona de desplazamientos visuales. El resultado global es una caída de la activación de la neurona de discrepancias por debajo de su tasa basal. La alteración de la activación de la neurona de discrepancias nos dice que el acontecimiento no se ha debido al movimiento de nuestro ojo, sino a alguna otra causa.
Para completar la historia, necesitamos que la neurona de discrepancias influya en nuestra impresión de si la escena ante nosotros es estacionaria o no. Esto está indicado por la flecha gris hacia abajo de la figura 61, que conecta la salida de la neurona de discrepancias con el sentido del desplazamiento visual. Si la neurona de discrepancias se activa a su tasa basal, entonces no se percibe ningún desplazamiento visual y la escena parece estacionaria, que es lo que ocurre cuando realizamos los movimientos oculares. Pero si la escena se desplaza por causas externas, entonces la neurona de discrepancias se activa de forma diferenciada por encima de su tasa basal, y percibimos un cambio externo. He aportado un ejemplo que utiliza los movimientos oculares hacia la derecha, pero el mismo principio valdría para los movimientos hacia la izquierda, hacia arriba o hacia abajo. En todos los casos aprendemos a calibrar nuestras acciones frente a los efectos que ocasionan y utilizamos entonces las discrepancias para inferir que se han producido acontecimientos externos.
Estos esquemas explican un efecto visual curioso: si a alguien se le paralizan los movimientos oculares con una dosis pequeña de curare, dirá que la escena que contempla tiende a agitarse a su alrededor cuando intenta mover los ojos[79]. Los ojos realmente no se mueven porque están paralizados, y sin embargo el paciente percibe el desplazamiento de su entorno. En estas situaciones, las neuronas de movimientos oculares están activadas, pero son incapaces de afectar a los músculos paralizados del ojo. Por lo tanto, las neuronas de movimientos oculares se activan sin un cambio en el impulso visual. El desequilibrio resultante hace que las neuronas de discrepancias se activen de modo diferencial por encima de su tasa basal, dándonos la sensación de que la escena se ha desplazado.
He ofrecido un esquema muy simplificado para resaltar que los principios con los que ya estamos familiarizados se pueden relacionar con el modo en el que se calibran los movimientos oculares frente a los cambios en el impulso visual que ocasionan. Todavía se conocen pocos detalles de los mecanismos neurales mediante los cuales se produce esta calibración. Se sabe que hay regiones del cerebro que están organizadas como mapas que relacionan los impulsos visuales con los movimientos del ojo, de modo similar a los mapas sensoriales y motores descritos en el capítulo 6[80]. Sin embargo, están menos claros los mecanismos por los que se modifica la fuerza de las sinapsis o su cantidad mediante la experiencia para asegurar que los movimientos oculares cuadren con los cambios en los impulsos visuales recibidos. Cualesquiera que sean los mecanismos, parece probable que estén basados en una realimentación del tipo descrito, en la cual las conexiones sinápticas se modifican gracias a las discrepancias entre las acciones y el efecto que se espera que produzcan.
§. Bucles de aprendizaje y acción
A menudo pensamos que la acción es algo que sigue a la percepción y el aprendizaje. Percibimos, aprendemos y entonces actuamos en consecuencia. Pero la calibración visual demuestra que la percepción, el aprendizaje y la acción están mucho más entrelazados[81]. Cuando realizamos una acción, como mover los ojos, lo que percibimos puede cambiar como resultado de esa acción, o sea, la escena vista se desplaza por la retina. Hay cambios sensoriales parecidos que también pueden deberse a acontecimientos externos. Una de las primeras cosas que el cerebro tiene que descifrar es la manera de diferenciar entre los cambios sensoriales ocasionados por nuestras propias acciones y los ocasionados por procesos externos. Si no lo hiciéramos, confundiríamos los movimientos de nuestro entorno con los resultados de nuestros propios movimientos. El problema se puede solucionar aprendiendo a calibrar nuestras acciones frente a los efectos que producen. Primero podemos aprender qué esperar de nuestras propias acciones. Si detectamos discrepancias respecto a las expectativas, podemos inferir que ha tenido lugar un cambio por causas externas.Tal y como se ilustró mediante nuestro esquema neural para los movimientos oculares, los principios implicados en este proceso de calibración no difieren de los que hemos tratado antes. Tenemos la interconexión del refuerzo y de la competencia en bucles de realimentación que intervienen en el funcionamiento de las neuronas de discrepancias. Se trata de bucles alimentados por un equilibrio de la variabilidad de la población y de la persistencia, ya que varias acciones y experiencias conducen a modificaciones duraderas en la fuerza de las sinapsis o en su cantidad. Y para que esto ocurra, los impulsos deben ocurrir muy próximos de modo que puedan cooperar. El resultado global es que aprenderemos a emparejar los efectos con las acciones que los generaron, y entonces podremos discriminar entre acontecimientos ocasionados por nuestra propia actividad y los que tienen un origen externo.
La calibración presenta otro beneficio: puede ayudar a guiar nuestras acciones. Tal y como se describió antes, si nos sentáramos en una habitación a oscuras y un punto luminoso saltara a una nueva posición, seríamos capaces de seguirlo con los ojos. De acuerdo con nuestro esquema neural, el movimiento inicial de la diana crea un desplazamiento visual que, a su vez, cambia la activación de la neurona de discrepancias (los ojos aún no se han movido). Cuanto más se desplaza la diana, más cambia la tasa de disparo de nuestra neurona de discrepancias, que de este modo nos informa de cuánto se movió la diana. Podemos utilizar esta información como auxilio para guiar el movimiento de los ojos. Para conseguirlo, la señal saliente de las neuronas de discrepancias se puede realimentar para mover los ojos. Los principios son los mismos que antes, salvo que ahora estamos utilizando las discrepancias no solo para detectar los acontecimientos externos sino que también realimentamos la información para afectar a nuestras acciones. Si la diana retrocediera disimuladamente durante las sacudidas oculares, el sistema automáticamente se recalibrará y se modificará en consecuencia el alcance de cada movimiento. La calibración no solo nos permite diferenciar los acontecimientos internos de los externos, sino que también ayuda a guiar nuestras acciones en respuesta al cambio externo.
§. Movimientos suaves
Hemos visto que la calibración se puede aplicar a los movimientos balísticos como las sacudidas oculares. Pero muchas de nuestras acciones son mucho más suaves que esta: no caminamos vacilantes como los robots, sino que disponemos de un conjunto de movimientos mucho más fluido. ¿Cómo aprendemos a controlar las acciones suaves?Las armas modernas son mucho más elaboradas que las de los tiempos de Leonardo. En vez de disparar un misil y confiar en que caiga donde queremos, podemos lanzar misiles teledirigidos que darán en la diana porque tienen sistemas sensoriales, como un radar integrado, que detectan los errores entre su trayectoria y la posición del objetivo, y ajustan su trayecto convenientemente. Los sistemas de control de esta clase tienen la ventaja de poder adaptarse si los misiles se desvían de su trayectoria por factores externos, como el viento; también tienen la flexibilidad necesaria para perseguir los objetivos en movimiento[82]. Nuestras acciones corporales cuentan con sistemas equivalentes. Como antes, quiero utilizar nuestra visión para ilustrar algunos de los principios involucrados[83].
Si miramos un objeto y giramos la cabeza lentamente, los ojos permanecerán fijos sobre el objeto por varias razones: una depende de neuronas en los ojos y en el cerebro que detectan la velocidad a la que se mueven las imágenes sobre la retina. Al igual que las neuronas de desplazamientos visuales que describí anteriormente, estas neuronas de la velocidad visual se dedican a comparar las señales que los diferentes fotorreceptores de la retina reciben a lo largo del tiempo[84]. Pero funcionan a una escala del tiempo más pequeña, mientras el ojo está detenido o se mueve lentamente, no entre saltos. A medida que se mueve la cabeza, las neuronas de la velocidad visual detectan la velocidad y la dirección con la que la imagen se mueve sobre la retina. Esta información se utiliza después para mover los ojos. Si la velocidad de los ojos coincide con la velocidad de la imagen detectada inicialmente, entonces la imagen vuelve a quedarse quieta sobre la retina y el objeto se mantiene visible. Pero si hay alguna discordancia, entonces la imagen comienza a deslizarse un poco por la retina, estimulando de nuevo a las neuronas de la velocidad visual, lo que provoca otro reajuste de la velocidad del ojo. De este modo, nuestros ojos utilizan un sistema de realimentación continua para seguir al objeto, de forma muy parecida a como los misiles guiados persiguen su objetivo. A diferencia de las sacudidas oculares, este tipo de movimiento ocular es suave, por lo que se puede fijar la mirada a medida que giramos la cabeza poco a poco.
Hay otro sistema que ayuda a mantener los ojos sobre el objetivo. Incluso cuando están cerrados, los ojos tienden a permanecer orientados hacia la misma posición externa mientras giramos la cabeza. Existe una tendencia natural a que roten en la dirección opuesta a la cabeza para contrarrestar el movimiento de la cabeza. Esta respuesta depende de los receptores neurales del oído interno que detectan el movimiento de la cabeza y envían la información para corregir el movimiento ocular. Al igual que con las neuronas de la velocidad visual, este sistema contrarresta el movimiento de la cabeza con un movimiento suave del ojo e impide que la imagen se deslice demasiado por la retina.
Tenemos dos sistemas que nos mantienen sobre el objetivo, uno percibe la velocidad visual y el otro percibe el movimiento de la cabeza. ¿Cómo se integran ambos sistemas? Como veremos, la respuesta es que uno se calibra con el otro.
En los años noventa del siglo XX, George M. Stratton llevó a cabo lo que Richard Gregory calificó como el experimento quizá más famoso de la psicología experimental[85]. Se puso durante varios días unas gafas que invertían la imagen vertical y lateralmente para averiguar si su vista se adaptaría a la nueva situación. Estas gafas invertían los movimientos de la cabeza, porque cuando giraba la cabeza hacia un lado, el campo visual iba en sentido opuesto a lo que cabría esperar. Cuando se colocó estas gafas por primera vez, Stratton se sintió muy confuso: cuando giraba la cabeza tenía la sensación visual opuesta a la que esperaba. Pero después de llevarlas continuamente durante una semana, encontró que las cosas volvían a parecer relativamente normales: se había adaptado a la nueva disposición. Los estudios posteriores demostraron que entre los efectos de tal adaptación estaba la inversión de la respuesta al movimiento de la cabeza: los ojos giran en la dirección opuesta a lo que harían normalmente cuando se gira la cabeza. Y ocurría incluso en la oscuridad. Aunque los ojos se movieran en la dirección opuesta, el efecto global era el mismo: reducir el deslizamiento de la imagen por la retina, aunque ahora con las gafas inversoras. ¿Cómo se produce este reajuste?
A medida que giramos la cabeza de izquierda a derecha, las neuronas de determinadas regiones del cerebro, como el cerebelo, aprenden a calibrar los impulsos desde el oído interno y desde la vista. Estas neuronas funcionarían como las neuronas de discrepancias que tratamos en las sacudidas oculares, y equilibran un impulso frente a otro. Después de la auto calibración, si giramos la cabeza con los ojos cerrados, se activan las neuronas de discrepancias en el cerebelo porque no hay ninguna señal visual que contrarreste el impulso desde el oído interno. Esta activación de las neuronas de discrepancias haría que los ojos giraran lo necesario. Después de llevar las gafas inversoras durante un tiempo, las neuronas de discrepancias se recalibran en la nueva situación al capturar la relación inversa entre las señales del oído interno y de las neuronas de la velocidad visual. El resultado es que, cuando se gira la cabeza, los ojos se impulsan en dirección opuesta a la de antes, lo que ayuda a mantener la mirada en la nueva situación.
Tal calibración, además de ayudar a mantener los ojos sobre el objetivo, ayuda a diferenciar entre los efectos de nuestros movimientos y los del entorno. Cuando George Stratton se puso por primera vez las gafas inversoras, tuvo la sensación de que se movía el exterior cuando movía la cabeza: «No me sentí como si estuviera mirando un conjunto de objetos quietos, sino como si todo lo que veía pasara rápidamente y oscilara ante mis ojos»[86]. La explicación es que su cerebro se había calibrado antes con la visión normal, y ahora estaba detectando las discrepancias entre sus acciones y los efectos visuales esperados. De acuerdo a nuestro esquema, se estaban activando determinadas neuronas de discrepancias, que estimulaban su sensación de movimiento externo. Después de llevar las gafas durante unos pocos días, se re calibraron sus neuronas de discrepancias, por lo que la sensación de movimiento que seguía a sus acciones había desaparecido.
La ventaja de auto calibrarnos de este modo es que seremos capaces de discriminar los acontecimientos externos de los movimientos ocasionados por nuestras propias acciones. Si miramos por la ventanilla de un tren en marcha, sentimos que el mundo se mueve respecto a nosotros. Las neuronas de discrepancias se pueden estar activando porque nos llegan señales de las neuronas de la velocidad visual, mientras que no hay ninguna señal que la contrarreste que venga de las neuronas que perciben o impulsan los movimientos de la cabeza[87].
La discrepancia resultante hace que sintamos que se mueve el exterior. Nuestra sensación de movimiento es relacional y se basa en que los impulsos sensoriales se desvían respecto a lo que nuestras propias acciones nos hacen esperar.
He puesto el énfasis en la manera en la que calibramos nuestras acciones frente a los impulsos visuales, pero lo mismo vale para los impulsos de los otros sentidos. Si permanecemos cerca de un hombre en una habitación a oscuras y nos estiramos para tocarlo, atribuiremos la percepción de la presión de nuestra mano a nuestra propia acción. El hombre está quieto y lo hemos tocado. Alternativamente, si mantenemos un brazo inmóvil y de repente sentimos que el cuerpo del hombre nos presiona la mano, suponemos que se ha movido él y no nosotros. La percepción de la mano puede ser exactamente la misma en ambos casos, pero solo en el segundo caso percibimos movimientos externos porque hemos aprendido a calibrar nuestras acciones corporales frente a los efectos que ocasionan. Mediante muchas acciones anteriores, comenzando con nuestros movimientos aleatorios cuando éramos bebés, determinadas neuronas de discrepancias pueden haber aprendido a calmarse hasta valores de actividad basal de acuerdo con los efectos de nuestros movimientos. Solo percibimos el movimiento externo cuando nuestras sensaciones se desvían de esta expectativa.
Todos estos tipos de calibración entran en funcionamiento cuando caminamos. A medida que paseamos por un paisaje quieto, sentimos que todo el movimiento tiene que ver con nuestras propias acciones. Al haber calibrado previamente las señales procedentes de nuestros pies, piernas, ojos y cabeza entre sí y con las neuronas que impulsan su actividad, somos conscientes de que andamos por un mundo fijo. Por el contrario, si nos paramos en un pasillo rodante, percibimos que nos están llevando. A pesar de que la información visual que recibimos es similar a la que recibimos al caminar, las neuronas que normalmente se activarían cuando movemos las piernas permanecen en silencio. Esta desviación de lo esperado puede estimular determinadas neuronas de discrepancias y dar la sensación de que el movimiento se impulsa desde el exterior. Si comenzamos a caminar por el pasillo rodante, entonces comenzamos a sentir que los desplazamientos visuales que percibimos son consecuencia de nuestras propias acciones. Pero todavía hay cierta discrepancia, porque los desplazamientos visuales son mayores de lo que esperaríamos según nuestras experiencias previas al caminar, y tendríamos la sensación de que andamos mucho más rápido de lo que solemos ir.
Tales calibraciones son importantes porque nos permiten afianzarnos en el mundo y actuar con más eficacia dentro de él. Si vemos que una manzana cuelga de un árbol, dirigirnos hacia la manzana y cogerla constituye una acción relativamente sencilla. Sin embargo, imaginemos que no hemos calibrado previamente nuestras acciones con sus efectos. A medida que miramos la manzana, la escena ante nosotros parecería que comienza a saltar a nuestro alrededor con cada movimiento ocular. Girar la cabeza hacia la manzana haría que el mundo girase sobre nosotros. Y a medida que intentáramos caminar hacia el árbol, sentiríamos que el suelo se mueve y que sube hasta encontrarse con el pie. Todo parecería dar vueltas a nuestro alrededor y cuanto más hiciésemos para corregirlo, peor lo estaríamos poniendo: en vez de caminar suavemente hacia la manzana, andaríamos dando traspiés como un borracho desconcertado.
La capacidad para sujetarnos al mundo depende de toda una serie de calibraciones cruzadas. Gracias a esta riqueza combinatoria, nuestras neuronas son capaces de integrar muchas acciones y sus efectos sensoriales. Esto nos permite aprender continuamente de las discrepancias al comparar un componente con relación a otro. Al aplicar de forma recurrente el mismo sistema relacional, aprendemos a calibrar muchos sistemas entre sí y a actuar con más eficacia. Los principios de la riqueza combinatoria y de la recurrencia no son solo importantes para aprender de nuestro entorno, sino que también son el centro de nuestras acciones.
§. Un periplo activo
En el capítulo anterior vimos que podíamos concebir el aprendizaje como un periplo por el espacio neural. Ahora vemos que este periplo no solo implica que respondamos a los acontecimientos que nos rodean, sino que actuamos continuamente, y con ello cambiamos lo que experimentamos. Cuando movemos los ojos o giramos la cabeza, vemos desde nuevas perspectivas. Podemos aprender del modo en el que nuestras acciones influyen en nuestra experiencia, y esta información se realimenta para influir en otras acciones y experiencias. Nuestro periplo por el espacio neural no está empujado pasivamente por el entorno y las percepciones, sino que interviene constantemente la realimentación con nuestras acciones.Al nacer ya contamos con algunas acciones y respuestas instintivas, y para darnos cuenta nos bastará con ver cómo un recién nacido mueve y tiende los brazos y las piernas. Estas reacciones instintivas proporcionan la rampa de lanzamiento para un viaje neural mucho más largo, durante el cual nuestras acciones y experiencias están de manera continua alimentándose mutuamente. Con este proceso aprendemos las relaciones entre las acciones y sus efectos, y esto nos conduce a nuevas regiones del espacio neural. La calibración ilustra cómo tiene lugar el aprendizaje. Pero este tipo de ejemplos no son más que la punta del iceberg.
Volvamos a los experimentos de Romo y Schultz del capítulo anterior, en los que el mono aprendía a responder al sonido de una puerta para luego recoger un trozo de manzana de una caja. Podemos dividir este proceso en dos tipos de aprendizaje: el primero consistiría en aprender la acción necesaria para retirar la recompensa (meter la mano en la caja); el segundo consistiría en aprender que el sonido de la puerta predice que esta acción será recompensada (sin el sonido de la puerta, la caja está cerrada e inaccesible para el mono). Ahora vamos a profundizar en cada una de estas etapas de aprendizaje.
Antes de que al mono se le enseñara a responder al sonido de la puerta, Romo y Schultz dejaron la puerta de la caja abierta durante un rato para permitir que el mono aprendiera que al meter la mano en la caja a veces le recompensaba con un trozo de fruta. Este aprendizaje surge a través del comportamiento exploratorio iniciado por el propio mono. El mono no necesita esperar una señal externa para explorar, sino que comienza con sus propios movimientos exploratorios dentro de un determinado contexto (en este caso, la jaula en la que está). Este comportamiento podríamos llamarlo curiosidad instintiva. Entonces, el mono encuentra que el inicio de algunas acciones, como meter la mano en la caja, a veces va seguido de recompensas. En otras palabras, aprende una acción relacionada con un objetivo, una expectativa de que se puede obtener una determinada recompensa después de iniciar una determinada acción. Esto se parece a lo que encontramos con la calibración visual, en donde se espera un desplazamiento visual determinado después de iniciar un movimiento ocular. La principal diferencia es que, para el aprendizaje exploratorio del mono, la relación entre la acción y el efecto es más complicada: solo algunas de las muchas acciones posibles obtienen una recompensa, y también varía el retraso entre el inicio de las acciones y sus efectos. No obstante, los principios básicos del aprendizaje a través de la interacción de acciones, expectativas y discrepancias son los mismos.
Al relacionar las acciones con sus efectos, el proceso del aprendizaje exploratorio no solo es similar a la calibración, sino que se construye sobre él. El mono es capaz de llevar a cabo movimientos exploratorios porque ya se ha sujetado al mundo y sabe cómo actuar dentro de él. Esta calibración precoz también significa que si la caja se mueve ligeramente, el mono sabe cómo ajustar sus movimientos para conseguir encontrar la recompensa. Es mediante la interacción entre la calibración y la exploración como se aprenden las acciones relacionadas con objetivos.
El entrenamiento con el sonido de la puerta introduce otro factor ambiental a este proceso. Ahora, el acceso a la caja está bloqueado a menos que suene la puerta. El mono aprende entonces que una determinada experiencia sensorial, que suene la puerta, significa que la acción de meter la mano en la caja tendrá una recompensa. Tal comportamiento puede surgir al extender el tipo de esquema de aprendizaje descrito en el capítulo anterior con la incorporación de acciones relacionadas con objetivos. En vez de limitarse a fortalecer las conexiones con determinados movimientos del mono, el impulso sensorial se acaba conectando con la acción relacionada con el objetivo de coger la recompensa. Esto significa que si la caja se mueve, el mono seguirá pudiendo realizar la acción apropiada.
Este ejemplo comporta una única acción relacionada con un objetivo, recoger la manzana de recompensa; pero también se pueden tener varios objetivos, lo que introduce más niveles de cooperación y de competencia. Supongamos que al mono se le presentan dos cajas con manzana de recompensa, una a la izquierda y la otra a la derecha, y que ambas se abren a la vez al sonar la puerta. Después de muchos intentos, el mono aprende que las recompensas se pueden encontrar en ambas cajas. Si el mono solo moviera un brazo, entonces tendría que elegir, cuando escuchara la puerta, entre meter la mano a la izquierda o a la derecha. Si la recompensa de cada caja es similar, entonces quizá se mueva con la misma probabilidad a la izquierda o a la derecha. Es importante que el mono tome una decisión, o de lo contrario se quedaría bloqueado, incapaz de decidir a qué lado ir y, por lo tanto, no conseguiría ninguna recompensa. Por lo tanto, las acciones posibles tienen que luchar entre sí en el cerebro para que una gane. Esta competencia puede transcurrir por caminos similares a los que sigue el desarrollo, en el que las células luchan por expresar un gen particular (Delta, como se explicó en el capítulo 3). En aquel caso, las células inhibían la actividad del gen en otra célula, con el resultado de que si la actividad de una célula era ligeramente superior a la de la otra, se reforzaba su ventaja y acababa por reprimir totalmente a la otra. La diferencia respecto a la decisión del mono es que la pugna tiene lugar a una escala de tiempo mucho más pequeña y las neuronas son las que inhibirán la activación de las otras. Por supuesto, los monos y los humanos no son conscientes de que se esté produciendo tal competencia neural cuando toman una decisión. Sentimos que tomamos decisiones por nuestra propia voluntad, pero entre bastidores las neuronas luchan hasta que al final prevalece una acción[88].
También tenemos la situación en la que cada caja contiene una recompensa diferente. Supongamos que las cajas a veces están vacías, y que quizá la caja de la izquierda está vacía más a menudo que la derecha. El mono se enfrenta ahora a una cierta incertidumbre sobre si conseguirá la recompensa o no. Con el tiempo, aprende a favorecer la caja derecha porque, de media, le recompensa más veces; pero también comprueba la caja izquierda con cierta frecuencia, por si acaso ha cambiado la probabilidad de encontrar alguna manzana. Está aprendiendo no solo a relacionar las recompensas con cada acción, sino también la mejor estrategia que debe adoptar para verse recompensado cuando se enfrenta a un abanico de elecciones y situaciones[89]. Tal aprendizaje de nuevo se conseguirá mediante la interacción entre las acciones, las expectativas y las discrepancias, aunque los esquemas neurales implicados sean más elaborados.
Otro factor que influye en tales elecciones es el incentivo, o valor relativo, asociado a las diferentes recompensas. Estos incentivos pueden a su vez cambiar según las experiencias del mono. Supongamos, por ejemplo, que el mono se ha estado alimentando con muchas manzanas y anhela una comida nueva, digamos un plátano[90]. Si ahora colocamos el plátano en la caja de la derecha, tras muchos ensayos, el mono aprende a ir a la derecha más a menudo que a la izquierda. Por el contrario, si el mono ya hubiera comido montones de plátanos, podría preferir ir a la izquierda para conseguir la manzana. El concepto de recompensa no es fijo, sino que también depende de la experiencia previa del mono.
El aprendizaje permite que el mono tome decisiones informadas sobre la mejor opción en función de su experiencia previa y la situación actual. Estas decisiones siempre se basan en relaciones, sopesando un conjunto de acciones frente a otro. Antes de que el plátano entrase en la escena, el mono seguramente se sentiría feliz al coger la manzana. Al introducir otra opción, se desplazan las expectativas y el comportamiento. Los principios básicos no son diferentes de los que nos hemos encontrado antes. En el centro del proceso se encuentran las neuronas de discrepancias, que funcionan de acuerdo con nuestros conocidos bucles gemelos de realimentación. Los bucles se alimentan por un equilibrio entre variabilidad (un abanico de experiencias y acciones) y persistencia (permanencia de la modificación en la fuerza y número de sinapsis). La principal diferencia es que hemos introducido más niveles de interacción entre las acciones y sus efectos, más grados de competencia, cooperación, riqueza combinatoria y recurrencia[91].
Sin esta profusa interacción entre la acción y la experiencia, nuestros periplos neurales serían muy diferentes. Supongamos que hubiésemos nacido sin la capacidad de movernos, pero con todos los sentidos intactos. Seríamos incapaces de mover los brazos o los ojos, o emitir un sonido, incluso aunque pudiéramos ver y escuchar lo que nos rodea (he supuesto que las funciones corporales como la respiración y el bombeo del corazón siguen funcionando para mantenernos vivos). En esta situación, tengo mis dudas de que fuéramos capaces de dar sentido a lo que escuchamos y vemos. Si alguien nos mostrara una manzana, tendríamos poca idea de lo que ha pasado, seríamos incapaces de seguirla con los ojos, y como nunca la habíamos cogido ni mordido, no tendríamos ni idea de su tacto ni su sabor. Incluso la noción de manzana como objeto externo nos resultaría extraña porque nunca habríamos tenido la ocasión de explorar activamente la relación entre nosotros y cualquier objeto de nuestro entorno. La imposibilidad de movernos sería una incapacidad mucho mayor que perder alguno de los sentidos. Un ciego o un sordo pueden todavía formarse una idea coherente del mundo porque son capaces de moverse por él y explorarlo con sus propias acciones. El psicólogo Hermann von Helmholtz resaltó la importancia de ser capaz de actuar para dar sentido al mundo, al escribir en 1866:
Solo conseguimos aprender a estar seguros de nuestros juicios sobre las causas de nuestras sensaciones cuando reunimos voluntariamente en diferentes relaciones los órganos sensoriales con los objetos. Esta clase de experimentación comienza en la más tierna infancia y continúa durante toda la vida sin interrupción. Si los objetos hubiesen simplemente pasado revista ante nuestros ojos por alguna fuerza extraña sin que pudiéramos interaccionar con ellos de ningún modo, probablemente nunca conseguiríamos orientarnos en tal fantasmagoría óptica.[92]
§. Se aprende con otros
Hasta ahora hemos estudiado que los individuos consiguen aprender gracias a sus propios esfuerzos. Pero existe otra opción, sobre todo para los animales sociales como los humanos: aprender de los otros. Podemos aprender a copiar las acciones de alguien o aprender de lo que hacen o dicen. Podría parecer que esta forma de aprendizaje es bastante diferente de lo que hemos descrito hasta ahora porque la información parece venir simplemente de fuentes externas. Sin embargo, incluso aquí, el aprendizaje puede estar cimentado en nuestras propias acciones.En los años noventa del siglo XX, Vittorio Gallese, Giacomo Rizzolatti y sus colaboradores en la Universidad de Parma estaban registrando las señales eléctricas de las neuronas cerebrales de los macacos[93]. Identificaron un conjunto determinado de neuronas que se activaban cuando un mono realizaba determinadas acciones relacionadas con objetivos, como coger fruta de una mesa. Cabe destacar que hallaron que estas mismas neuronas también se activaban cuando el mono observaba al experimentador u otro mono realizar la misma acción. Aquí, el objetivo es importante: si un mono observa que alguien agarra algo sin que haya ninguna fruta en la mesa, entonces no se activan las neuronas. Se las denomina neuronas especulares porque responden a las acciones externas de un modo que replica las acciones impulsadas por el propio animal.
Las neuronas especulares indican que nuestra respuesta a los otros estaría estrechamente conectada con nuestras propias acciones. A medida que aprendemos acciones relacionadas con objetivos, podemos fortalecer las conexiones neurales entre determinadas acciones y sus efectos. El mono es testigo de los movimientos de su brazo a medida que lo extiende para alcanzar la fruta, y los asocia con la obtención de una recompensa. Cuando el mono ve que otro individuo realiza el mismo tipo de acción, entonces se podrían estimular algunas de estas mismas neuronas. El resultado global es que las acciones de otros individuos se conectan a las acciones del propio mono, conexión que le permitiría interpretar lo que otros individuos están haciendo y, por lo tanto, aprender de ellos. El aprendizaje relacionado con conocer o imitar a los otros no es un fenómeno unidireccional, sino que está cimentado en el modo en el que aprendemos de nuestras propias acciones y en el que utilizamos los conocimientos para ayudar a forjar relaciones con los otros. Del mismo modo que una persona inmovilizada no comprendería que tiene una manzana ante sí, tampoco se harían una idea de lo que significa que alguien agarre dicha manzana: al no haber realizado nunca tal movimiento por sí mismos, la acción les resultaría absolutamente misteriosa.
Esta interacción entre nuestros movimientos y los de otros da cabida a otro tipo de cooperación. Si estamos hablando con alguien, realizamos toda clase de acciones: además de hablar una parte del tiempo, movemos los ojos, cambiamos la expresión de la cara y corregimos la postura. El interlocutor percibe estas acciones y le estimulan determinadas reacciones: sus ojos podrían seguir a los nuestros y su expresión facial podría cambiar según lo que digamos y hagamos. A su vez, nosotros percibimos estas reacciones, por lo que nuestras acciones nos vuelven por mediación del interlocutor. Por supuesto, al interlocutor también le ocurre lo mismo: ve reflejadas sus acciones a través de nosotros. Tenemos dos bucles de realimentación engarzados en los cuales nuestros movimientos y los de nuestro interlocutor están entrelazados. Esta cooperación mutua permite que nuestro cerebro se comunique con eficacia con el suyo. Lo vemos más claro cuando se rompe uno de los bucles de realimentación: si los ojos del interlocutor pierden brillo, o si responde de un modo que demuestra que no ha escuchado lo que decíamos, sentimos que la comunicación se ha bloqueado. Nuestras acciones ya no reciben la realimentación esperada del interlocutor y se pierde la intimidad. Como dijo el neurocientífico Chris Frith: «Cuando dos personas interaccionan cara a cara, intercambian los significados mediante un proyecto cooperativo»[94].
Los senderos neurales que tomamos al llegar al mundo no son independientes de los tomados por otros. Nuestros periplos influyen y responden continuamente a los senderos que nos rodean. Los principios básicos mediante los cuales esto acaba sucediendo pueden no ser muy diferentes de los que ya hemos descrito. En el centro de este proceso se encuentra la detección y el aprendizaje a partir de discrepancias mediante nuestro ya conocido doble bucle de realimentación. La única diferencia que ahora afecta a la interacción entre nuestras acciones y experiencias, y las de otros, es que hay más niveles de cooperación, de riqueza combinatoria y de recurrencia.
Hemos visto que la receta creativa para la vida proporciona un marco de trabajo conceptual para conocer nuestra capacidad de aprendizaje a partir de nuestras propias acciones y de las de otros. Incluso así, coger una fruta o tener una conversación podría parecer muy alejado de la aparición de una nueva idea o de la creación de un cuadro hermoso. ¿Qué hay detrás de esa inteligencia y creatividad, y cómo se relaciona con los procesos que hemos tratado? Para responder esta pregunta, estudiaremos cómo aprendemos a ver las cosas de determinada manera.
Capítulo 9
Visión por comparación
§. El ojo neuralDiez años después de que Cézanne pintara el retrato del marchante de arte Ambroise Vollard (lámina 1), Pablo Picasso pintó su propia versión del mismo tema (figura 62, lámina 2). De estilo cubista, el retrato de Picasso es muy diferente del de Cézanne: mientras que este último retrata su modelo como una forma sólida con bloques de color, Picasso utiliza elementos angulares y una estrategia más abstracta. Al explorar de diferente manera cómo ven lo mismo, cada artista captura diferentes aspectos y destaca cada relación a su manera.
§. Varios ojos
§. Vemos mediante modelos
§. Aprendizaje a muchos niveles
§. Ascendente y descendente
§. Interpretaciones en pugna
§. Una cuestión de estilo
§. Actos creativos
Apreciar las relaciones desde perspectivas nuevas no es solo una característica del arte, sino que reside en el centro de todas las formas de creatividad humana. Wordsworth vio sus propias andanzas solitarias en las de una nube. Newton relacionó la caída de una manzana con la órbita de los planetas alrededor del sol. Beethoven exploró nuevas relaciones tonales cuando compuso sus sinfonías. A una escala más modesta, realizamos actos creativos equivalentes cada día, mediante actividades como ordenar las flores en un jarrón o construir una nueva frase cuando hablamos[95]. En todos estos casos, las relaciones se exploran de nuevas maneras, sin importar que se trate de objetos, movimientos, sonidos o palabras.
En los dos capítulos anteriores examinamos cómo aprendemos a predecir y a actuar. Ahora quiero estudiar cómo aprendemos a interpretar o a ver las cosas que nos rodean de un modo particular. Veremos que estos tres procesos (predicción, acción e interpretación) están íntimamente conectados, gracias a lo cual llegaremos a las raíces de la creatividad humana. Tal y como nos podríamos imaginar, estas raíces no necesitan ningún principio fundamentalmente nuevo, sino que encontraremos otra aplicación de nuestra conocida receta creativa. Pero en la receta intervendrán otras ramificaciones relacionadas con el modo en el que las neuronas interaccionan y compiten unas con otras.

Figura 62. Retrato de Ambroise Vollard, Pablo Picasso, 1909. Véase la lámina 2.
Nuestras percepciones también adolecen de ciertas constricciones. Cuando contemplamos cuadros ambiguos como el del centro de la figura 63, solo hacemos una interpretación de cada vez (o vemos una anciana de perfil o una joven dando la espalda), pero nunca ambas al mismo tiempo. Nuestro cerebro parece obligado a resolver el cuadro de un modo o del otro. De igual forma, lo que vemos puede estar influido por lo que ya tenemos en mente. Si miramos primero el cuadro de la izquierda de la figura 63 y luego pasamos a mirar el central, es más probable que veamos a la anciana. Por el contrario, si miramos la imagen central después de haber contemplado la de la derecha, es más probable que veamos a la joven. Lo que vemos podría depender de lo que hayamos visto antes o de lo que esperamos ver. Estas expectativas están enraizadas en experiencias aún más antiguas. Si nunca hubiéramos visto una anciana o una joven, veríamos el panel central de la figura 63 con una luz diferente. Quizá lo veríamos como un patrón abstracto en blanco y negro, que sería otra interpretación válida. Nuestras interpretaciones no vienen dadas, sino que están constreñidas por la historia de nuestras experiencias.

Figura 63. Dibujo central: Mi mujer y mi suegra. Edwin Boring, 1930. El dibujo se ha modificado para resaltar a la anciana (izquierda) y a la joven (derecha).
Cuando intentamos atravesar un territorio prácticamente desconocido, una estrategia consiste en encaminarse en una dirección determinada para hacerse una idea de su configuración. La dirección elegida quizá no sea la mejor, pero al seguirla se nos revelarán algunos de los problemas a los que nos enfrentaremos. Esta es la estrategia que voy a seguir en este capítulo: quiero presentar una manera concreta de considerar la base neural de las interpretaciones. Mi objetivo no consiste en proporcionar una explicación definitiva porque, sin duda, muchos de los detalles que aparecerán serán incorrectos, sino que deseo seguir este camino para resaltar algunos principios fundamentales y los problemas que le afectan. El camino puede resultar arduo a veces, pero recordemos que lo importante no son los pormenores del viaje, sino lo que aprendemos durante el camino.
Los ejemplos se basarán principalmente en la interpretación de las imágenes visuales, aunque los principios que se tratarán valdrían igualmente para otras modalidades sensoriales, como el oído o el tacto. Como siempre, la simplificación inicial del problema nos ayudará a abordarlo: en vez de tratar con la interpretación de imágenes complejas, como una cara, quiero comenzar con algunos casos unidimensionales más simples.
Supongamos que vivimos en un mundo que contiene personas lineales. Cada persona estará formada por una línea gris clara con una región oscura dentro. La figura 64 muestra dos tipos de personas que podríamos encontrarnos. Una de ellas es María, reconocible porque la región oscura está centrada. La otra es Juan, cuya región oscura está ligeramente descentrada. Cuando miramos la figura 64, vemos que Juan es como María, pero con la región oscura desplazada ligeramente a la derecha. Somos capaces de ver esta relación porque estamos muy acostumbrados a mirar e interpretar los objetos que nos rodean. El problema consiste en concebir un sistema neural que consiga el mismo resultado, o sea, un sistema que vea a Juan y a María como la misma clase de entidad, al mismo tiempo que también detecte que son diferentes. Para ayudar con este problema, voy a introducir lo que denomino ojos neurales, que aunque en principio parezcan más bien teóricos, finalmente veremos que se corresponden con diferentes niveles de procesamiento en el cerebro.
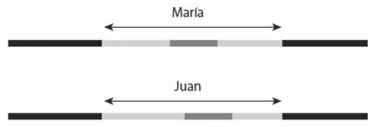
Figura 64. Representación unidimensional de Juan y María.
§. El ojo neural
Cuando movemos los ojos, el cambio de su posición produce el cambio correspondiente de los impulsos neurales que llegan a nuestro cerebro. Pero se puede conseguir lo mismo sin mover el ojo, solo con los elementos básicos que se ilustran en la figura 65. A la izquierda de esta figura se ve una neurona (la neurona receptora) que recibe los impulsos estimuladores de una neurona situada a su izquierda (I) y de otra situada a su derecha (D). La misma disposición se repite a la derecha de la figura, pero añadiendo una neurona más que denomino neurona de selecciones. La activación de la neurona de selecciones bloquea en la neurona I su capacidad para enviar señales a la neurona receptora, como si el cable de conexión se hubiese roto. Esto puede ocurrir mediante un proceso denominado inhibición pre sináptica, en el cual la activación de una neurona interfiere con la capacidad de otra para liberar neurotransmisores desde su terminación[98]. El resultado es que la neurona receptora solo tiene una conexión viva con la neurona D, mientras que la conexión con la neurona I está muerta (se muestra en gris). La neurona de selecciones ha hecho cambiar lo que el receptor está escuchando. Este cambio es reversible: si la neurona de selecciones deja de estar activada, entonces la conexión a la neurona I revive y el receptor vuelve a escuchar a las neuronas I y D.En este ejemplo, la activación de la neurona de selecciones simplemente bloquea la transmisión de una línea de comunicación. Se pueden concebir esquemas ligeramente más complicados, en los que la activación de una neurona de selecciones puede desplazar la conexión viva de una neurona a otra. Por ejemplo, supongamos inicialmente que la conexión de la izquierda está viva y la de la derecha inhibida, y que la activación de una neurona de selecciones destruye la conexión izquierda al mismo tiempo que se libera la de la derecha. En este caso, la activación de la neurona de selecciones traslada con eficacia la atención del receptor desde una neurona a la otra. De nuevo, el cambio es reversible: si la neurona de selecciones deja de estar activada, la conexión viva vuelve a ser la del impulso desde la izquierda.
Si aplicamos esta idea a muchas neuronas, llegaremos a la situación ilustrada en la figura 66: el panel superior muestra la imagen de María sobre algunos fotorreceptores retinianos. Estas neuronas envían su señal a otro conjunto de neuronas, que funcionan en lo que denomino el nivel 1. He mostrado dos conexiones entre cada neurona de nivel 1 y los fotorreceptores de la retina, una de la cuales está viva (en negro) mientras que la otra está muerta (en gris). Las conexiones vivas apuntan directamente a la retina. Debido a la disposición ordenada de estas conexiones, el nivel 1 replica esencialmente el patrón de actividad de la retina, que forma lo que se conoce como un mapa retino tópico. Las conexiones grises están ordenadas de un modo parecido, pero están desfasadas y forman conexiones diagonales con los fotorreceptores retinianos. Aunque muestre conexiones directas entre el nivel 1 y la retina, las conexiones pueden ser mucho más indirectas y combinarse en diferentes niveles intermedios (más tarde veremos que el nivel 1 puede corresponder a una región de nivel superior en la corteza visual del cerebro). Lo realmente importante para lo que vendrá después es que las diferentes conexiones con el nivel 1 están dispuestas de manera retino tópica, en fiel reflejo de la organización del ojo.

Figura 65. Selección de los impulsos que llegan a una neurona
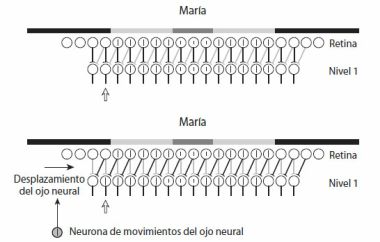
Figura 66. Observación de María con las conexiones verticales vivas de nivel 1 (panel superior), o después de un movimiento del ojo neural tras el cual las conexiones diagonales vuelven a estar en funcionamiento (panel inferior). La longitud de las líneas verticales dentro de las neuronas indica el nivel de actividad neural.
Supongamos que ahora aprendemos a seguir a María con nuestro ojo neural a medida que se mueve respecto a nosotros, a la izquierda o a la derecha sobre la línea negra de fondo (este movimiento también se puede generar cuando nos movemos respecto a ella). Esto implicaría una forma de calibración similar a lo que encontramos en el capítulo anterior: detectaríamos discrepancias cuando se mueve respecto a nosotros y, entonces, ajustaríamos nuestros ojos neurales para reducirlas. Sin embargo, ahora estamos calibrando nuestros movimientos del ojo neural en vez de los movimientos físicos. Calibramos de acuerdo con una perspectiva particular de María: cuando está en el centro. Nuestro ojo neural siempre intenta conservar esta imagen original a medida que María se traslada.
Esta calibración requiere que nuestro sistema neural sea capaz de detectar desplazamientos de la posición de María. Recordemos que ser capaz de detectar los desplazamientos visuales o el movimiento requiere combinar la información de varios fotorreceptores. En la figura 67 se muestra un posible esquema de la manera en que nuestro ojo neural podría hacerlo, en el cual he introducido un nivel más (nivel 2) que contiene tres neuronas, A, B y C (ABC), que reciben los impulsos desde el nivel 1. La neurona A mira el tercio izquierdo del campo visual, la neurona B el tercio central y la neurona C el tercio derecho. Para simplificar las cosas, he mostrado solo las conexiones funcionales entre la retina y el nivel 1, no las muchas conexiones inactivas (grises) que también están presentes. A medida que María se traslada ligeramente de izquierda a derecha sobre el fondo, podemos aprender a seguir su movimiento mediante los movimientos del ojo neural, que siempre intenta mantener la perspectiva original que tenemos de ella. Los cambios detectados por las neuronas ABC regresarían a la neurona de movimientos del ojo neural para que este le siga la pista. El resultado neto es que si María se mueve respecto a nosotros, el ojo neural la seguirá para mantenerla a la vista, lo que tiene el efecto de estabilizar la imagen de María, al menos para las regiones del cerebro que funcionan de este modo.
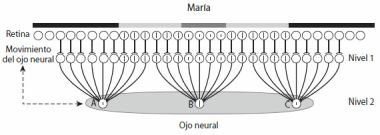
Figura 67. Conexión de los movimientos del ojo neural con las señales visuales recibidas por las neuronas ABC.
§. Varios ojos
Hasta ahora, el movimiento de un ojo neural no ha conseguido nada que no pudiéramos hacer ya físicamente moviendo un ojo real. Pero los ojos neurales tienen una gran ventaja sobre los normales: podemos tener más de dos. Tal y como apuntaron David Van Essen y colaboradores en Caltech en 1993, la interpretación visual se conseguiría gracias a la existencia de muchos ojos neurales[99]. Para hacernos una idea de cómo podría funcionar esto, tenemos que añadir otro nivel a nuestro esquema.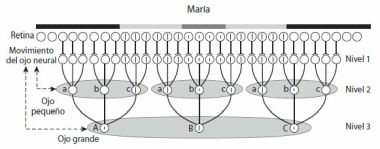
Figura 68. Ojos neurales con un nivel más.
A medida que nuestros ojos neurales siguen los movimientos de María, emergen ciertas tendencias. Los tres ojos pequeños, por ejemplo, siempre tienden a moverse juntos. Si María se mueve a la derecha, los tres ojos neurales también lo hacen y muestran correlaciones en los patrones de su movimiento. Nuestro sistema neural podría aprender estas tendencias mediante la alimentación de los impulsos desde las neuronas de movimientos del ojo neural a lo que yo llamo neuronas de correlaciones, que pueden capturar las principales maneras en que los impulsos tienden a activarse juntos. No necesitamos entrar en los detalles del mecanismo porque intervienen bucles neurales de aprendizaje y principios similares a los encontrados antes para las neuronas de discrepancias[101]. A medida que nuestros ojos neurales siguen a María, la fuerza de las sinapsis entre las neuronas de movimientos del ojo neural y las neuronas de correlaciones se modifican poco a poco, por lo que las neuronas de correlaciones comienzan a responder de una manera que es consecuencia de que los ojos neurales tiendan a moverse juntos. A través de este aprendizaje, nuestro sistema neural espera este patrón de movimientos del ojo neural, y cualquier desviación de él se registraría como una discrepancia. Habiendo entrenado nuestro sistema neural sobre María, ¿qué ocurre cuando miramos a Juan? Ya que los ojos neurales han aprendido a mantener una perspectiva particular de María, intentarán hacer lo mismo con Juan. En la figura 69 se muestra un posible resultado. El ojo grande y los ojos pequeños de la izquierda y de la derecha están en las mismas posiciones que antes, pero el ojo pequeño central ha desplazado su punto de vista hacia la derecha para conseguir que la región oscura esté a la vista. Al desplazar su mirada, el ojo neural central ahora recibe la misma entrada que si estuviera mirando a María, por lo que la señal visual que entra en cada ojo neural es exactamente la misma que cuando los ojos neurales estaban mirando a María. Estamos mirando a Juan, pero lo vemos como veríamos normalmente a María. Estamos viendo a Juan como María.
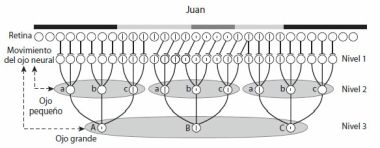
Figura 69. Cómo se mira a Juan después de haber entrenado los ojos neurales sobre María
El sistema neural ha logrado hacer lo que nos habíamos propuesto que hiciera. Los ojos neurales nos dicen que Juan y María son la misma clase de cosa (vemos a Juan como María), pero nuestras neuronas de correlaciones también nos dicen que hay una diferencia entre ellos; Juan y María desencadenan diferentes patrones de activación en las neuronas de correlaciones. He explicado este ejemplo en detalle para resaltar algunas características generales de cómo aprendemos a interpretar lo que vemos. Veamos ahora algunas de las peculiaridades.
§. Vemos mediante modelos
Cuando miramos a María, nuestro sistema neural construye una expectativa, o un modelo, de lo que ve, lo que implica que el sistema aprende lo que cada ojo neural detecta normalmente y adónde tienden a ir los diferentes ojos neurales. He presentado un ejemplo unidimensional simple, pero lo mismo valdría para los objetos más complejos que vemos a nuestro alrededor. Cuando miramos a alguien, su apariencia puede cambiar de todas las maneras posibles: puede girar la cabeza, mover la boca o cambiar de expresión. Estas alteraciones son mucho más complicadas que el movimiento de María de izquierda a derecha. Sin embargo, al igual que con los movimientos de María, hay elementos que vemos que se mueven de manera correlacionada. Cuando giramos la cabeza, los ojos se mueven con ella; cuando alguien sonríe, la izquierda y la derecha de la boca se mueven al unísono. De algún modo nuestro cerebro hace un seguimiento de estos acontecimientos, lo que nos permite desarrollar expectativas o modelos de la manera en que los diferentes elementos de la cara tienden a moverse al unísono.Para formar un modelo de esta manera, aprovechamos algo que se suele dar por sentado: la continuidad física. Suponemos que frente a nosotros siempre está la misma persona mientras la observamos. Cuando miramos continuamente a María durante un tiempo, confiamos en que no nos la han cambiado por otra en dicho intervalo. Esta suposición nos permite construir un modelo para esa persona, una representación de cómo se organizan sus diferentes elementos. Si los objetos saltaran continuamente de una identidad a otra, sería muy difícil que estableciéramos expectativas o modelos. Si un objeto cambiara de parecerse a una manzana a parecerse a un plátano, o a una cara, y luego a un árbol, nos resultaría muy difícil formarnos una imagen de él. En la práctica, los objetos no muestran tal comportamiento errático, por lo que somos capaces de aprender lo que se denominan representaciones invariables, o sea, modelos que nos permiten percibir una identidad invariable cuando estamos mirando algo[102]. Conservamos este concepto sin referencia a las variaciones ocasionadas por los cambios en una pose o expresión, del mismo modo que retenemos la misma imagen neural de María a medida que se traslada. Al hacerlo, podemos también aprender la manera en que los diferentes elementos de nuestro objeto se comportan unos respecto a otros. Aprendemos las correlaciones que intervienen cuando una cara se vuelve o sonríe. Se mantiene la identidad a pesar de que cambien algunos elementos.
Una vez que hemos establecido un modelo o expectativa para un objeto, podríamos ser capaces de ver objetos relacionados a través del mismo modelo. Veremos a Juan a través del modelo que nos formamos cuando observamos a María, lo que nos permite ver una cosa como versión de otra: vemos a Juan como María. Pero de todas formas somos conscientes de que no tenemos exactamente lo mismo frente a nosotros debido a las discrepancias con el modelo que nos habíamos construido. Hay un aspecto doble en relación con la visión por comparación. Tenemos un modelo neural común a través del cual vemos los objetos relacionados, pero nuestro sistema neural también registra las discrepancias con el modelo, lo que nos permite saber que dos objetos son diferentes.
A medida que vemos el objeto nuevo durante un rato, podemos comenzar a aprender el patrón de las discrepancias neurales que detectamos. Lo que distingue a Juan, lo que se vio inicialmente como una discrepancia, se convierte en una expectativa si comenzamos a ver a Juan con regularidad. Podemos capturar esta expectativa mediante los circuitos de aprendizaje en los que intervienen otras neuronas de correlaciones, y llegar a un modelo más amplio que incluye a Juan y a María. A este modelo más amplio podríamos llamarlo un modelo de persona. Cuando miramos a Juan o María mediante el modelo de persona, cada uno desencadena un patrón concreto de activación de neuronas de discrepancias y de correlaciones, lo que nos permite identificar de quién se trata. Si vemos una tercera persona que no se parece a Juan ni a María, detectaríamos discrepancias con nuestro modelo de persona, lo que nos conduce a otras identificaciones y elaboraciones. Estamos continuamente construyendo correlación tras correlación para establecer modelos más generales y elaborados, aprendiendo cada vez del patrón de discrepancias.
Este proceso de construcción de modelo sobre modelo forma parte del periplo neural que cada uno de nosotros sigue desde el momento en que nacemos. Todos comenzamos al nacer con determinadas expectativas (modelos) incorporadas en las conexiones neurales y en su distinta fuerza. También nacemos con unas disposiciones neurales estructuradas que permiten que elaboremos y construyamos estos modelos mediante el aprendizaje. Para nuestro esquema neural, estas disposiciones estructuradas serían los diferentes ojos neurales y el modo en el que se comportan juntos. En consecuencia, modificamos nuestros modelos con la experiencia, y aprendemos las diferentes correlaciones a partir del modo en el que el mundo cambia a medida que lo vemos y, entonces, utilizamos esta información para elaborar aún más nuestros modelos. Se trata del principio de recurrencia aplicado a los modelos neurales.
En el centro de este proceso de construcción de modelos se encuentra el aprendizaje a través de las discrepancias y de las correlaciones. Como vimos con los esquemas neurales para las neuronas de discrepancias en los dos capítulos anteriores, este aprendizaje se basa en nuestros conocidos bucles dobles de realimentación entre el refuerzo y la competencia, alimentado por un equilibrio entre variación y persistencia. Basándose en una población de experiencias (variación), determinadas neuronas potencian su propia fuerza sináptica (refuerzo) de un modo que también genera sus propias limitaciones (competencia), y esto conduce a cambios duraderos de la fuerza de las sinapsis (persistencia). Los mismos principios se aplican a los esquemas de aprendizaje para las neuronas de correlaciones, aunque no hayamos entrado en detalles. Los bucles de realimentación son más complicados porque ahora tenemos un número mayor de neuronas de discrepancias y de correlaciones que funcionan en combinación, pero los principios centrales son exactamente los que hemos tratado antes.
§. Aprendizaje a muchos niveles
Otra característica clave de nuestro esquema neural es que en él intervienen interacciones entre varios niveles. No teníamos un solo ojo neural, sino varios, cada uno a un nivel concreto. Estos ojos neurales nos permitían ir más allá de lo que conseguirían los ojos normales. Mientras que un ojo normal solo puede apuntar en una única dirección en un momento dado, nuestros muchos ojos neurales pueden apuntar en varias direcciones al mismo tiempo, lo que nos permite aprender sobre muchos aspectos de un objeto al mismo tiempo. Esto es lo que nos permitía ver a Juan como María, pues el ojo pequeño neural del centro podía mirar a la derecha mientras los otros ojos miraban de frente.Los diferentes niveles de nuestro esquema no trabajan por separado, sino de una manera muy cooperativa. Lo que ve el ojo neural grande depende de hacia dónde estén mirando los ojos pequeños neurales, porque la señal visual tiene que atravesarlos para alcanzar el ojo neural grande. Y a la inversa, lo que ven los ojos pequeños neurales depende del ojo neural grande porque su movimiento hace que todos los impulsos también se desplacen. Esta relación recíproca permite que los ojos se ayuden mutuamente. Por ejemplo, una limitación de los ojos pequeños es que de forma individual no consiguen detectar grandes cambios debido a que su campo visual es pequeño. Un gran desplazamiento podría sacar el objeto de su margen, y sería un movimiento que capturaría con más facilidad el ojo grande, al tener un alcance más amplio porque capta toda la escena. Al seguir un gran movimiento, el ojo grande devolvería con eficacia el objeto a la vista de los ojos pequeños, quienes entonces detectarán cualquier otro cambio más pequeño. Y a la inversa, el ojo grande no tiene flexibilidad para seguir los detalles de la imagen, mientras que los ojos pequeños sí pueden y, por lo tanto, ayudarán a ofrecer una imagen más coherente al ojo grande. La cooperación dentro de los niveles y entre ellos resulta decisiva en tales sistemas.
En nuestro esquema simplificado teníamos una retina con unos 30 fotorreceptores y tres niveles, pero podríamos imaginarnos una retina con muchos más fotorreceptores y más niveles. La retina humana, por ejemplo, tiene cerca de 100 millones de fotorreceptores, por lo que hay muchas señales que podrían penetrar en muchos niveles de integración. Si añadimos otro nivel a nuestro esquema, cada uno de nuestros tres ojos pequeños neurales tendría sus propios tres ojos neurales que lo alimentan, por lo que nuestro objeto acabaría recibiendo las miradas de trece (9 + 3 + 1) ojos neurales. Con dos niveles más tendríamos 121 (81 + 27 + 9 + 3 + 1) ojos neurales, es decir, podríamos mirar en 121 diferentes direcciones al mismo tiempo, y cada una captaría aspectos de un objeto a una escala concreta.
Realmente, en el cerebro humano la información visual se trata a muchos niveles[103] , algunos de los cuales se muestran en la figura 70. Las señales procedentes de los fotorreceptores de la retina pasan primero a las neuronas del interior del ojo, denominadas células del ganglio retiniano. Las señales emitidas por los ganglios retinianos van a una región del cerebro denominada cuerpo geniculado lateral o CGL, que envía impulsos a la región V1, que a su vez envía impulsos a una región denominada V2. Esta a su vez está conectada a otras regiones, como V 3 y V4. Todas estas regiones (los ganglios retinianos, el CGL y las V1, V2, V3 y V 4) están organizadas de manera retinotópica: todas tienen un patrón espacial de sensibilidades que es reflejo del de los fotorreceptores del ojo. Su información visual también se combina de diferente forma. Los campos receptivos de las neuronas de V4 tienden a ser más grandes que los de V2, que a su vez tienden a ser más grandes que los de V1.
Estas regiones cerebrales se sabe que interaccionan de diferentes maneras entre sí y con otras áreas que intervienen en la visión. Sin embargo, todavía está poco claro cómo, gracias a estas interacciones, se aprende de las interpretaciones visuales concretas. El esquema que he presentado basado en los movimientos de los ojos neurales es un modo de concebir lo que podría ocurrir. El nivel 1 de mi esquema está relacionado con una de las áreas visuales del cerebro. Los movimientos del ojo neural podrían corresponder a desplazamientos en los impulsos que alcanzan esta región visual. Los requisitos clave son que el área que corresponda al nivel 1 esté organizada retinotópicamente y que haya más niveles por encima de ella que integren las señales que emite y devuelva información para modificar sus impulsos de entrada. Tanto si en la formación de modelos visuales interviene algo similar al esquema que he presentado de los ojos neurales o algún otro mecanismo, tener varios niveles que cooperen resultará probablemente decisivo. Imaginemos lo que ocurriría si miráramos las cosas a través de un único nivel. Si únicamente pudiéramos captar la imagen muy detallada, adquiriríamos muchísima información, pero no sabríamos cómo integrarla. Algo así les ocurre a las personas que padecen agnosia visual de formas[104]: cuando se les presenta un objeto, pueden ver muchos detalles visuales, pero no son capaces de reconocer el propio objeto. Por el contrario, si solo pudiéramos captar la información visual en el nivel más amplio, obtendríamos una perspectiva global, pero al carecer de información detallada, solo tendríamos una idea vaga de lo que estaríamos viendo. Gracias a que vemos a través de varios niveles de cooperación conseguimos llegar a interpretaciones y reconocimientos concretos.
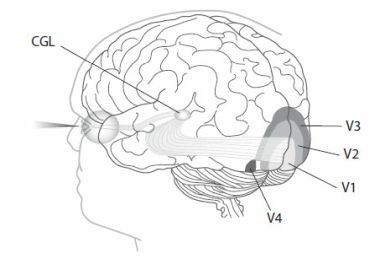
Figura 70. Algunas áreas del cerebro implicadas en el procesamiento visual.
§. Ascendente y descendente
Cuando miramos a nuestro alrededor, es tentador pensar que lo que vemos se basa en una cadena de eventos de procesamiento. Recibimos información por los ojos, y la procesamos mediante una serie de etapas en el cerebro para que nos informe de lo que estamos mirando. Esto a veces se llama una perspectiva ascendente de la percepción porque comienza con los simples sentidos y termina con las funciones superiores del cerebro. En cambio, la perspectiva descendente consiste en acercarse al mundo a través de modelos concretos que obtenemos de nosotros mismos y no de nuestros sentidos, lo que nos permite ver e interpretar la información sensorial que recibimos.Nuestro esquema neural ilustra de qué modo estas dos perspectivas, ascendente y descendente, no son realmente excluyentes, sino que están entrelazadas. Cuando vemos a María, los impulsos visuales viajan desde los fotorreceptores de la retina a los diferentes ojos neurales de nivel superior, así que las señales van viajando en sentido ascendente. Los cambios detectados en los ojos neurales de los niveles 2 y 3 sirven para realimentar las conexiones con el nivel 1, lo que influye en la dirección en la que miran los ojos neurales[105]. Tenemos un flujo de información descendente que, a su vez, cambia las señales que llegan a los ojos neurales de nivel superior desde el nivel 1. Tenemos una realimentación continua entre los diferentes niveles, y no un flujo de información unidireccional.
De igual forma, cuando miramos a Juan, el modelo que hemos aprendido al observar a María se utiliza para dirigir los ojos neurales. Se trata de un proceso descendente porque vemos a Juan a través del modelo de María que llevamos implantado. Pero a medida que miramos a Juan durante un rato, nuestras neuronas de correlaciones aprenden las discrepancias concretas y elaboran un modelo de persona más amplio. Este proceso implica el aprendizaje ascendente porque las señales sensoriales influyen ahora en el modelo. De nuevo se da una realimentación continua entre los modelos y la información sensorial que detectan. La interpretación no depende únicamente de nosotros, ni únicamente de nuestras entradas sensoriales, sino de la continua competencia entre las dos en el tiempo.
En el capítulo anterior vimos que nuestro periplo neural implica un diálogo continuo bidireccional entre lo que hacemos y lo que experimentamos. Las interpretaciones también se pueden explicar con una interacción bidireccional parecida. Al igual que aprendemos sobre la forma en que nuestras acciones físicas se relacionan con las experiencias, aprendemos que las acciones internas, simbolizadas como movimientos del ojo neural, se relacionan con los objetos que detectan. A través de la realimentación continua entre la actividad neural interna y la información sensorial llegamos a modelos o interpretaciones concretos que, a su vez, guían otras interacciones y experiencias, y a una reelaboración de nuestros modelos. Nuestro periplo por el espacio neural no es ni ascendente ni descendente, sino una mezcla inseparable de los dos.
§. Interpretaciones en pugna
Volvamos sobre el problema de por qué alternamos la interpretación de los cuadros ambiguos, como el de la anciana y la joven. Supongamos que después de haber aprendido un modelo de persona para María y Juan, nos presentan una imagen que combina las características de ambos, tal y como se muestra en la parte inferior de la figura 71. El extremo izquierdo de la región oscura se alinea con María, pero el derecho lo hace con Juan. Nuestros ojos neurales y las neuronas de correlaciones pueden estar repartidas entre dos opciones: verlo como María o como Juan. El ojo pequeño neural del centro, por ejemplo, podría dibujarse hacia la derecha o hacia el centro, o quizá hacia un punto entre ambos. ¿Qué ocurre en esta situación? Una posibilidad es que nuestro sistema neural no tome partido: no ve el objeto ni como María ni como Juan. Otra opción es que se decante por uno de ellos, y decida que quizá sea María. Puede mantenerse fiel a su decisión, o cambiar de vez en cuando, con lo que a veces lo verá como María y otras veces como Juan. En cualquier caso se toma alguna decisión en lugar de deambular continuamente por tierra de nadie.
Figura 71. Los unidimensionales María y Juan, y una persona ambigua.
¿Por qué están estructurados nuestros sistemas neurales para resolver asuntos en vez de merodear por los estados intermedios? La respuesta se basa, en primer lugar, en el motivo por el que realizamos interpretaciones. Supongamos que María es una buena amiga, mientras que Juan a menudo nos resulta desagradable. Las expectativas sobre la persona que vemos dependerá entonces de si pensamos que es María o Juan. Si se trata de María, podríamos predecir una conversación agradable, mientras que si se trata de Juan, podríamos anticipar que el encuentro nos resultará menos agradable. Esto, a su vez, influye en nuestras acciones, pues resultará más probable que nos paremos a hablar con María, mientras que evitaremos a Juan en la medida de lo posible. Al final tendremos que decidir qué acción tomar, hablar o evitar, por lo que es importante resolver de qué persona se trata. Por lo tanto, tiene sentido que nuestros sistemas neurales estén estructurados para suministrar determinadas interpretaciones en vez de titubear continuamente sin ofrecer ninguna resolución. La razón fundamental por la que hacemos interpretaciones es ayudar a guiar nuestras acciones, por lo que es importante que podamos resolverlas.
Para ver lo críticas que resultan tales resoluciones, imaginemos que vamos por la vida sin ser capaces de realizar interpretaciones decisivas. Podríamos dudar si debemos sentarnos en lo que parece una silla porque en otros aspectos se parece también a una mesa. Nos preguntaríamos si deberíamos dar un mordisco a una manzana, porque también se parece en cierto modo a una pelota de tenis. O quizá dudemos si abrir una puerta, porque también se parece a una ventana. La vida sería imposible si dejáramos sin resolver nuestras interpretaciones de un modo u otro. Sin estas resoluciones, seríamos incapaces de actuar, quedaríamos atrapados en un mar de dudas. Este problema se soluciona haciendo que las interpretaciones compitan entre sí: mediante la inhibición neural cruzada, las interpretaciones se pueden volver mutuamente excluyentes, por lo que tenderemos a clasificar o separar nuestras experiencias de un modo u otro. Tal clasificación nos ayuda a actuar de un modo particular, a decidir que nos comeremos una manzana porque no se presta a confusión con una pelota de tenis, o a atravesar una puerta porque es diferente de una ventana. Nuestras categorías no permanecen inalterables, sino que las modificamos mediante el aprendizaje. Sin embargo, al crear cierto tipo de clasificación, seremos capaces de ordenar nuestro entorno con eficacia. Tal y como vimos al considerar el desarrollo biológico, ordenamos el mundo ordenándonos a nosotros mismos. Pero en vez de ordenarnos mediante las interacciones intercelulares inhibidoras que definen las regiones del embrión, ahora lo hacemos mediante neuronas que se inhiben unas a otras según nuestras experiencias después de nacer.
Nuestras interpretaciones dependen de los mismos principios que nos hemos ido encontrando en este libro. El doble bucle de realimentación de refuerzo y competencia anda detrás del aprendizaje mediante discrepancias y correlaciones, como se ilustró con los esquemas neurales para las neuronas de discrepancias en los dos capítulos anteriores. Estos bucles gemelos son alimentados por nuestra colección de experiencias y acciones, y conducen a cambios persistentes en la fuerza de las sinapsis y en su cantidad. Los bucles también están impregnados con diferentes formas de cooperación, competencia, riqueza combinatoria y recurrencia neurales. Existe cooperación entre varios niveles, simbolizados por los ojos neurales que actúan en combinación para producir numerosos modelos o interpretaciones de lo que vemos. Las interpretaciones también compiten entre sí para que veamos un objeto de un modo u otro. Todo esto ocurre al desplazar recurrentemente las discrepancias y las expectativas. La complejidad de nuestras interpretaciones no surge de la introducción de nuevos principios, sino de proporcionar más ramificaciones al modo en el que funcionan nuestros bien conocidos principios.
§. Una cuestión de estilo
Para demostrar que los mismos principios pueden conducir a interpretaciones muy elaboradas, quiero continuar perfeccionando nuestro esquema de ojos neurales y aplicarlo a imágenes más complejas. La meta consiste en capturar una de nuestras interpretaciones más escurridizas, a saber, el estilo de un artista.En nuestro ejemplo simplificado de personas unidimensionales teníamos tres niveles de ojos neurales que aprendían a seguir aspectos concretos de una persona, como la región oscura central. Ahora imaginemos que se extiende este esquema a imágenes bidimensionales y a varios niveles neurales, quizá con cientos o miles de ojos neurales en el equivalente del nivel 2. A medida que vemos objetos complejos, como una cara, durante cierto tiempo, nuestros ojos neurales aprenden a seguir algunos aspectos a distinta escala. Un ojo neural grande podría seguir la cara en su conjunto, mientras que un ojo neural pequeño que mire hacia el centro podría seguir la punta de la nariz, o uno que mire más abajo podría seguir la barbilla. Esta idea está ilustrada en la figura 72, que muestra dónde están mirando algunos de estos ojos neurales pequeños cuando observamos los retratos de Rembrandt ( izquierda) o Modigliani (derecha). He indicado las posiciones observadas por los ojos neurales con puntos sobre las peculiaridades, como el centro de la barbilla, el extremo izquierdo de la boca, etc. Cada punto está colocado en una relación fija respecto a los otros (punto blanco en el ojo izquierdo, negro en el ojo derecho, etc.). Fui capaz de colocar los puntos de esta manera porque ya sabía cómo había que mirar e interpretar las imágenes de rostros en dos dimensiones. Encontré con facilidad las peculiaridades correspondientes, como la barbilla o el ojo izquierdo, en ambos retratos. Pero imaginemos que estas posiciones fueran localizables automáticamente por cada ojo neural debido al modo en el que han sido entrenados gracias a la exposición a muchas caras en el pasado. De hecho, he conseguido identificar las peculiaridades, como los ojos y la nariz, con tanta eficacia porque mi propio cerebro ya se había expuesto antes a este entrenamiento.

Figura 72. Posiciones en las que se fijan siete ojos neurales en sendos cuadros de Rembrandt y de Modigliani. Izquierda: detalle de Retrato de Maria Trip, Rembrandt van Rijn, 1639.Derecha: detalle de Retrato de una joven, Amedeo Modigliani, 1917-1918.
El retrato medio representa una expectativa, o modelo, de las posiciones a las que tenderán a acudir nuestros ojos neurales. Cuando miramos cualquier retrato, los ojos neurales no van precisamente a estas posiciones medias, sino que se desvían un poco. Los ojos neurales en los retratos de Rembrandt y Modigliani de la figura 72, por ejemplo, no están exactamente en las mismas posiciones que en el retrato promediado. Cada retrato, por lo tanto, representa una desviación de nuestra media, una discrepancia de nuestras expectativas. Las discrepancias no son aleatorias, sino que muestran ciertas tendencias. En un retrato de alguien con la cabeza girada hacia la izquierda, muchos de nuestros ojos neurales se desplazarán juntos en la misma dirección. O si el retrato es de alguien que sonríe, los ojos neurales que rodean la boca se mueven hacia arriba de un modo coordinado, y capturamos estas tendencias con las neuronas de correlaciones. Gracias al refuerzo, a la competencia y a la cooperación, estas neuronas destilan las diferentes tendencias, la manera en la que nuestros ojos neurales tienden a desviarse de su posición media, tal y como la vemos en la población de 179 retratos.

Figura 73. Retrato promediado, en el que se superponen las posiciones medias en las que se fijan los ojos neurales (izquierda). Los 179 retratos utilizados para construir las medias son de Rembrandt, Modigliani, Leonardo, Velázquez, Soutine, Giotto, Duccio, Freud y una selección de retratos funerarios romanos.
La neurona de correlaciones del panel inferior de la figura 74 responde de un modo diferente porque se activa con frecuencia ante retratos con una gran nariz y un cuello largo que no llevan sombrero, pero lo hace muy poco frente a retratos con las características opuestas. La he denominado neurona de cabezas largas y destocadas. En nuestra colección de retratos también los hay con nariz y cuello más largos (como los de Modigliani) que no llevan sombrero. Esta correlación la captura la neurona de cabezas largas y destocadas.
Cuando vemos un determinado retrato, nuestras neuronas de correlaciones se activarán a un ritmo determinado. Un retrato de Modigliani con una cabeza delgada larga y sin sombrero, por ejemplo, haría que tanto las neuronas de cabeza estrechilarga como las de cabeza alargada larga y destocada se activaran con bastante frecuencia. He mostrado los patrones de activación que desencadenan algunos retratos de Rembrandt y Modigliani en la figura 75. En este diagrama, la tasa de activación de la neurona de cabezas estrechilargas forma el eje horizontal, mientras que la tasa de activación de la neurona de cabeza alargada y destocada forma el eje vertical. La combinación particular de niveles de activación desencadenada por cada retrato se puede entonces representar mediante el punto en el que se cortan los niveles de activación. Estas posiciones (calculadas por el ordenador) se muestran como puntos gris claro en los retratos de Rembrandt y gris oscuro en los retratos de Modigliani. Observemos que los de Modigliani tienden a concentrarse alrededor de la parte superior derecha del diagrama, porque la mayoría de ellos tienen cabezas estrechilargas sin sombrero. Por lo tanto, desencadenan una fuerte activación de la neurona de cabezas estrechilargas y de la neurona de cabeza alargada y destocada. El Retrato de Jeanne Hébuterne de Modigliani, por ejemplo, activa con frecuencia tanto la neurona de cabezas estrechilargas como la neurona de cabeza alargada y destocada, colocándola en el extremo superior a la derecha. No obstante, se dan excepciones: el retrato de Oscar Miestchaninoff de Modigliani, por ejemplo, está ubicado cerca del centro en vez de en la parte superior derecha porque no todos los retratos de Modigliani tienen cabezas estrechilargas: sus retratos de hombres, en particular, tienden a tener caras mucho más redondeadas y cuellos más cortos, colocándolos entre los de Rembrandt.
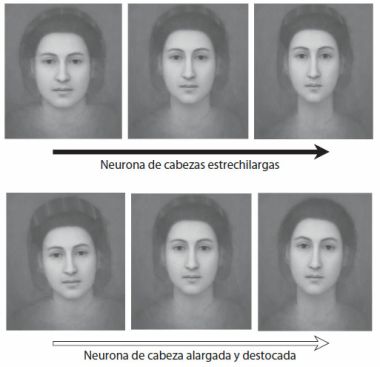
Figura 74. Dos de las tendencias capturadas por las neuronas de correlaciones de la colección de 179 retratos. Véase la lámina 10.

Figura 75. Patrones de activación de las neuronas de correlaciones desencadenadas por los retratos de Rembrandt (puntos gris claro) y de Modigliani (puntos gris oscuro). El cruce en el centro muestra la posición del retrato promediado. Se muestran dos retratos distintos: derecha: Retrato de Jeanne Hébuterne, de Amedeo Modigliani, 1919. Arriba: Oscar Miestchaninoff, de Amedeo Modigliani, 1917.
Figura 76. Tendencia capturada por la neurona de Modigliani.
Figura 77. Esbozos que corresponden a la activación frecuente de la neurona de Modigliani (derecha) en comparación con el esquema del retrato promediado (izquierda).
El ejemplo que he ofrecido sobre la extracción de tendencias de una selección de retratos a través de las interacciones neurales es muy artificial y simplificado. Solo he descrito unas pocas tendencias de una pequeña colección de retratos. De igual forma, solo he tenido en cuenta unos pocos ojos neurales y he ignorado algunos aspectos de los cuadros, como el contraste o el color. En la práctica, podemos aprender de muchos más patrones neurales de los que he indicado, lo que nos permitirá ser mucho más eficaces a la hora de discernir el estilo. Esta es la razón por la que nos damos cuenta de que el cuadro de Modigliani Oscar Miestchaninoff es diferente de los retratos de Rembrandt, aunque se solape con el agrupamiento de Rembrandt en nuestro gráfico. En cualquier caso, los elementos básicos del reconocimiento estilístico no serían diferentes de lo que he descrito para la neurona de Modigliani.
Para llegar a la neurona de Modigliani no me ha hecho falta introducir ningún principio fundamental nuevo, solo más grados de interacción en nuestro esquema neural. La neurona de Modigliani no es diferente de las otras, sino que adquiere sus propiedades por meras interacciones con otras neuronas y no por la adición de algún otro ingrediente. El aspecto del estilo capturado por la neurona Modigliani no es una peculiaridad de la neurona aislada, sino el resultado del efecto combinado de un gran número de interacciones neurales ascendentes y descendentes. Además, la señal emitida por la neurona de Modigliani puede a su vez volver a influir en estas interacciones. Después de que gracias al aprendizaje se haya consolidado una neurona de Modigliani, se verá estimulada cuando observemos a una mujer con una cara y un cuello largos, que nos recordará a un retrato de Modigliani. Hasta cierto punto, vemos a la persona como un cuadro de Modigliani, mientras que también somos conscientes de que estamos viendo a una persona y no un cuadro. La estimulación de la neurona de Modigliani no es solo un resultado, sino que realimenta nuestro sistema neural e influye en cómo vemos las cosas.
Aunque resulte difícil definir un estilo artístico, los mecanismos neurales subyacentes no necesitan ser tan especiales: simplemente son el reflejo de que nuestra conocida receta creativa actúa a medida que interaccionan entre sí muchos niveles neurales, modelos y experiencias.
§. Actos creativos
Imaginemos a Cézanne pintando sobre un lienzo. Después de colocar algunas áreas iniciales de color, puede sentir que algunas cumplen sus expectativas, mientras que otras no son realmente correctas o crean efectos asombrosos. Podría entonces reaccionar a esta mezcolanza de expectativas y discrepancias añadiendo más pinceladas, elaborando el cuadro e incitando nuevas valoraciones y comparaciones. El cuadro progresa así como si fuera un diálogo crítico entre Cézanne y lo que ve ante sí. A veces quizá sea consciente de este diálogo, pero las más de las veces este se producirá de forma inconsciente: se sentirá impulsado instintivamente a añadir algo de color aquí o allí, o sentirá que una pincelada es la correcta o que no pega[109]. Este diálogo creativo implica una interacción compleja entre todos los procesos que hemos ido tratando (predicciones, acciones e interpretaciones). Cézanne no se limitaba a mezclar y colocar colores al azar, sino que basándose en su experiencia como pintor, predecía la clase de efecto que probablemente tendrían, y utilizaba este conocimiento para guiar sus actos. Entonces interpretaba los colores que había colocado en el lienzo, exploraba sus relaciones y las comparaba con el motivo que estaba pintando. Las concordancias y las discrepancias que iba encontrando le conducirían a más predicciones y acciones, con lo que el proceso continuaba avanzando. El cuadro surgía gracias al diálogo neural que entrelazaba las predicciones, las acciones y las interpretaciones a muchos niveles. Cada vez que pintaba un cuadro, Cézanne también seguramente incrementaba su habilidad, modificaba sus modelos neurales de tal modo que la próxima vez que se planteaba un cuadro, lo pintaba de un modo ligeramente diferente. El resultado no era solo un cuadro, era la propulsión del cerebro de Cézanne a través del espacio neural.Lo mismo se puede decir de cualquier acto creativo: cuando un poeta compone un poema, hay un diálogo neural continuo en su cerebro en el que siguen interviniendo secuencias verbales. A diferencia de la pintura, no hay que realizar ninguna acción física explícita, pues el poeta no necesita poner nada por escrito ni hablar en voz alta mientras compone. Pero tal como vimos al tratar de los ojos neurales, hay una delgada línea entre las acciones físicas y las internas. Un movimiento de los ojos neurales puede tener las mismas consecuencias sobre la percepción que un movimiento físico del ojo, aun cuando el primero implique un ajuste interno de la actividad neural (cuyas conexiones están en funcionamiento), mientras que en el segundo interviene un movimiento físico explícito. La única diferencia entre el poeta y el pintor es que la obra del primero puede restringirse a algo meramente interno, sin que implique necesariamente acciones físicas ni efectos sensoriales. Esto no significa que el poeta trabaje en un vacío sensorial, puesto que el lenguaje que ha utilizado lo aprendió mediante interacciones anteriores con otros y con el entorno. Incluso si compone un poema a oscuras en una habitación silenciosa, lo construye sobre un periplo neural de gran alcance durante el cual el poeta aprendió muchas relaciones entre las predicciones sensoriales, las acciones y las interpretaciones.
Lo mismo ocurre con la creatividad de un científico: su cerebro también juega con las expectativas y las discrepancias, basándose en el aprendizaje y experiencia. Gracias a este diálogo podrá resolver determinadas discrepancias, que le conducirán a nuevos modelos o modos de conocimiento. A su vez, estos pueden conducir a acciones concretas, como la realización de un experimento para evaluar un modelo. A continuación, los resultados experimentales pueden realimentarse para influir en las siguientes etapas del periplo neural, lo que conducirá a otros refinamientos del modelo. Al igual que con los pintores y con los poetas, una gran parte de la creatividad de un científico se produce de forma inconsciente, sin que tenga conciencia plena de lo que está pasando. Al científico se le puede ocurrir de repente una nueva explicación, igual que un poeta puede pensar inesperadamente un verso[110]. Darwin dejó constancia del placer que sintió cuando de repente se dio cuenta de que la divergencia de las especies estaba relacionada con la diversidad del entorno: «Puedo recordar el punto exacto del camino, mientras iba en el carruaje, en el que, para mi gozo, se me ocurrió la solución»[111]. No cabe duda de que este problema le preocupó durante mucho tiempo, en el que las discrepancias iban y venían por su cerebro, lo que finalmente provocó una modificación de sus modelos neurales que redujo determinadas discrepancias y dio lugar a la repentina comprensión que sintió en el carruaje.
Los actos creativos de artistas, poetas y científicos, o de alguien que simplemente dispone unas flores en un jarrón, se pueden ver como producto de los procesos que ya hemos tratado, o sea, implican una interacción recurrente entre predicción, acción e interpretación. Hemos visto que cada uno de estos procesos depende del aprendizaje mediante discrepancias y correlaciones, los cuales, a su vez, están dando vueltas en nuestro doble bucle de realimentación de refuerzo y competencia, impulsado por un equilibrio entre la variabilidad de la población y la persistencia. Los actos creativos no requieren ningún nuevo ingrediente ni principio especiales, sino tan solo más niveles de cooperación, interacción combinatoria y recurrencia. Se pueden ver como una manifestación de la receta creativa para la vida.
Esto puede parecer una visión algo empobrecida de la creatividad. ¿Cómo es que algo tan maravilloso como nuestra propia capacidad para crear puede reducirse a una receta de interacciones cuyos fundamentos no difieren de los que encontramos en otras transformaciones de los seres vivos? Con seguridad, la creatividad humana presenta cualidades diferentes a las de un perro que está aprendiendo a salivar con el sonido de una campana, o a las de un mono que mete la mano para conseguir la manzana de recompensa. Creo que tal juicio cualitativo no es más válido que decir que un sapo es cualitativamente diferente del huevo del que procede, o que la bacteria nadadora E. coli es cualitativamente diferente de los primeros organismos vivos que aparecieron hace unos pocos miles de millones de años. Estas diferencias que nos parecen cualitativas no surgieron gracias a ningún elemento misterioso nuevo, sino a consecuencia del modo en el que funcionan e interaccionan los ingredientes de la receta creativa para la vida.
Desde Darwin, los científicos han intentado trazar una línea clara entre la creatividad y las transformaciones del mundo biológico para conseguir derrumbar la falacia de que los procesos vivos se explican mediante un principio rector externo o creador. Pero esta separación ha tenido el triste efecto de desconectar la creatividad humana de los otros procesos vivos, al colocarla en una categoría especial que nos separa de nuestros parientes animales. Aunque esta separación pueda resultarles cómoda a algunos, creo que es un punto de vista equivocado y empobrecedor. En mi opinión, si apreciamos las similitudes formales entre nuestros actos creativos y los otros procesos de los seres vivos, obtendremos una perspectiva más enriquecedora de la vida en su conjunto, y un punto de vista más apropiado del lugar que ocupamos en ella. Esto no disminuye lo maravillosa que es la creatividad humana, sino que la coloca en un contexto biológico más amplio.
Hasta ahora, he resaltado las similitudes entre el aprendizaje, la evolución y el desarrollo, que se pueden ver como manifestaciones de la misma receta creativa. Pero hay otra conexión entre ellas: del mismo modo que vimos que el desarrollo se cimienta en la evolución (capítulo 5), el aprendizaje está impregnado de la evolución y del desarrollo. Nuestros tres ejemplos de la receta creativa para la vida están conectados por la historia y por la forma. Ahora quiero estudiar más de cerca esta relación histórica.
Capítulo 10
Encuadremos las recetas
§. Desarrollo del aprendizajeLa historia de Simbad el Marino comienza en una calle estrecha y concurrida de Bagdad. Un día, cuando un porteador caminaba por la calle llevando una pesada carga sobre la cabeza, decidió sentarse para descansar en el exterior de una de las casas. El porteador escuchó cantar y olió una comida deliciosa que venía de la casa y del jardín, lo que le hizo exclamar lo injusto que era que alguna gente disfrutara tanto con tan poco esfuerzo, mientras que él sufría llevando cargas pesadas a cambio de una pequeña remuneración. Apenas había acabado de decir esto, cuando un criado salió de la casa y le dijo al porteador que su amo quería hablar con él. El amo, Simbad, se dirigió al porteador con amabilidad, le hizo sitio en el banquete, y comenzó a relatar al porteador y a los otros invitados la historia de sus muchas aventuras y largos viajes.
§. Instintos básicos
§. Flexibilidad frente a rigidez
Esta introducción a Simbad el Marino se conoce como un marco narrativo. Presenta el contexto para los cuentos que contará Simbad, y proporciona una narración que enmarca a otras narraciones que se cuentan en ella. El marco narrativo del porteador está a su vez enmarcado por otra historia, pues Simbad el Marino es uno de los cuentos de Las mil y una noches, un libro que contiene los muchos cuentos que la joven Sherezade contó a un rey al que tenía que entretener con una historia cada noche para impedir que la ejecutara. Uno de los cuentos es el del porteador y Simbad.
Estas historias contadas en distintos niveles comparten algunas similitudes. Todas se cuentan para entretener a una determinada audiencia: Las mil y una noches está escrito para entretener a sus lectores, Sherezade entretiene al rey y Simbad entretiene a sus huéspedes. Las historias también se rematan al final con un final feliz. Las mil y una noches termina con la boda del rey con Sherezade, al final de la historia de Simbad, el porteador se da cuenta de que Simbad realmente se había ganado su situación, y la historia de Simbad termina con su regreso final a Bagdad. Las diferentes historias tienen una forma común además de que unas enmarcan a las otras.
Un tipo similar de relación se aplica a la evolución, al desarrollo y al aprendizaje. Su forma común es que se basan en la receta creativa para la vida. En sus relaciones de encuadre intervienen sus orígenes históricos. La evolución encuadra al desarrollo porque mediante la evolución surge el desarrollo. Y tanto la evolución como el desarrollo enmarcan al aprendizaje, porque gracias a estas transformaciones previas se hace posible el aprendizaje. Ya hemos estudiado la relación entre la evolución y el desarrollo (capítulo 5). En este capítulo quiero estudiar la forma en que la evolución y el desarrollo enmarcan al aprendizaje.
§. Desarrollo del aprendizaje
El aprendizaje no comienza con una pizarra en blanco, sino que siempre está basado en un contexto neural concreto. El aprendizaje DT descrito en el capítulo 7, por ejemplo, requiere determinadas conexiones entre las neuronas que se dedican a detectar recompensas, discrepancias, expectativas e impulsos sensoriales. De igual forma, la calibración y la capacidad para formar determinadas interpretaciones dependen de las organizaciones neurales que ya existan. Estas conexiones y relaciones iniciales se remontan al proceso del desarrollo. A través del desarrollo de los embriones, los animales nacen con cerebros, cuerpos y sistemas sensoriales muy estructurados que van a necesitar para el aprendizaje.No obstante, la transición entre el desarrollo y el aprendizaje no es neta: a diferencia de la narración de Simbad, que comienza con una frase concreta, la transición entre el desarrollo y el aprendizaje es mucho más difusa. La activación de las neuronas no comienza con el nacimiento, sino que ya tiene lugar durante el desarrollo embrionario. El desarrollo no se detiene al nacer, sino que continúa a medida que el recién nacido crece y madura. Una buena ilustración de la interacción sutil entre el desarrollo y el aprendizaje procede de algunos estudios sobre la vista.
En el capítulo anterior vimos que en la parte posterior del cerebro había una región relacionada con la vista, que se denomina corteza visual primaria o V1. Si apuntamos con un haz de luz sobre una pantalla enfrente de un gato o mono anestesiados, solo una pequeña región de V1 mostrará una fuerte actividad eléctrica. A medida que movemos la luz de un lugar a otro en el campo visual para estimular una parte diferente de la retina, también se mueve la región de actividad eléctrica en V1. Es como si hubiera un mapa en V1 conectado con un mapa correspondiente en la retina. Se dice que la región V1 del cerebro está organizada como un mapa retinotópico.
Los animales suelen tener dos ojos, por lo que hay dos posibles mapas retinotópicos en V1, uno para cada ojo. ¿Cómo están integrados estos mapas? En los años sesenta del siglo XX, David Hubel y Torsten Wiesel, que trabajaban en la Harvard Medical School de Boston, comenzaron a tratar esta cuestión mediante una serie de registros detallados de la corteza visual de gatos y monos[112]. Al igual que nosotros, en estos animales se solapan los campos visuales del ojo izquierdo y del derecho. Hubel y Wiesel encontraron que cuando se muestra una pequeña barra de luz en una posición sobre una pantalla, primero a un ojo y luego al otro, se activa una región similar de V1 en ambos casos. Al mover la barra a una nueva posición se desplaza la región activada en V1 para ambos ojos en paralelo. Por lo tanto, las conexiones procedentes de ambos ojos están, en líneas generales, alineadas entre sí en V1.
Sin embargo, también encontraron un patrón más fino: dentro de una región concreta de V1 observaron que algunas neuronas respondían principalmente a la luz que incidía en el ojo izquierdo, mientras que otras reaccionaban principalmente a la luz que incidía en el derecho. Los impulsos procedentes de los dos ojos se mantenían en cierta medida separados, aunque se mapearan sobre la misma región de V1. Por fin se dieron cuenta de que en realidad se trataba de dos mapas estrechamente entrelazados en V1, una versión izquierda y una versión derecha. La relación entre estos mapas de V1 del mono macaco se muestra en la figura 78. Las regiones negras muestran dónde domina un ojo, digamos el derecho, mientras que las regiones blancas muestran dónde domina el otro, el izquierdo. Es como si hubiera dos copias del mapa visual, uno para cada ojo, que se intercalan ligeramente entre sí, produciendo un mosaico de impulsos alternados de izquierda y derecha, conocido como patrón de dominancia ocular. ¿Cómo se establece este patrón?
Hubel y Wiesel decidieron estudiar lo que ocurriría si se privaba a los animales de la visión con uno de los ojos o con ambos mientras crecían. Los gatitos abren los ojos por primera vez de diez a doce días después de nacer. Para impedir que pudiera ver con un ojo, digamos el ojo izquierdo, Hubel y Wiesel cosieron quirúrgicamente los párpados de ese ojo de un gatito recién nacido, lo que le impedía recibir por él cualquier impulso visual. Cuando abrieron quirúrgicamente el ojo izquierdo unos pocos meses después, encontraron que el gatito había perdido irreversiblemente la vista en ese ojo. En vez de responder a ambos ojos, la V1 solo tenía fuertes conexiones con el ojo derecho, el único que había permanecido abierto. La primera hipótesis fue que las conexiones sinápticas del ojo cerrado se debilitaron simplemente a consecuencia de la falta de uso. Para comprobar esta idea, repitieron el experimento, pero ahora cerraron quirúrgicamente ambos ojos después del nacimiento, esperando que la vista de ambos ojos quedaría intensamente atrofiada. Sorprendentemente, encontraron que las conexiones no estaban tan mal, como si cerrar un segundo ojo hubiera ayudado al primero. No era tan solo la falta de uso lo que ocasionaba una ceguera después de privar temporalmente la vista, sino que había cierta clase de interacción entre ambos ojos. La interacción se describe mejor si volvemos al patrón de dominancia ocular de la figura 78: cuando se cierra quirúrgicamente un ojo, en vez de tener un patrón de dominancia ocular con bandas alternantes de la misma anchura, las bandas del ojo activo se hacen mucho más anchas. Un conjunto de bandas, digamos las bandas negras de la figura 78, se expanden y dejan solo tiras blancas muy finas que corresponden al ojo cerrado. Esto significa que el gato mantiene muchas conexiones neurales con el ojo abierto y relativamente pocas con el cerrado. Cuando se cierran ambos ojos, ninguno toma el control, por lo que el patrón de bandas final sigue teniendo unas anchuras similares para ambos ojos, como en un gato normal. Parece que la anchura de las bandas depende de una competencia entre los ojos: si uno es más activo, toma el control del espacio que normalmente formaría conexiones con el otro. Si ambos son igualmente activos o igualmente inactivos, las conexiones se comparten con más equidad.

Figura 78. Patrones de dominancia ocular en la corteza visual primaria del mono macaco.
No obstante, el patrón de bandas no depende únicamente de la experiencia visual. Como se mencionó anteriormente, si ambos ojos se mantienen cerrados desde el nacimiento, se sigue formando un patrón de dominancia ocular en la corteza visual, aunque comienza a desaparecer después de unas cuatro semanas. Esta tendencia instintiva a formar un patrón de bandas depende del proceso de desarrollo, y en ella intervendría la actividad neural espontánea en determinadas regiones del cerebro. En vez de existir una línea nítida entre el desarrollo y los cambios neurales ocasionados por las experiencias sensoriales, existe una interacción estrecha entre los dos: el desarrollo conduce a patrones concretos de organización neural; entonces, las experiencias modulan estos patrones y construyen sobre ellos, lo que permite que las relaciones entre las neuronas queden vinculadas de una forma más precisa con lo que experimenta el animal.
Esta transición difusa entre el desarrollo y el aprendizaje se debe a que los dos procesos comparten muchos mecanismos celulares y moleculares. Tal y como vimos en el capítulo 6, los cambios a corto y largo plazo en la fuerza sináptica implican la señalización molecular, el crecimiento o la muerte de las sinapsis, y el encendido o apagado de determinados genes. Los componentes moleculares del aprendizaje no son diferentes de los que intervienen en el desarrollo. Sin embargo, con el aprendizaje se integran de un modo concreto que les permite capturar las interacciones con el entorno a través de la modificación de rutas neuronales. Mediante este proceso de modificación, los periplos neurales que comienzan en el útero son empujados hacia nuevas direcciones, de acuerdo con lo que se aprende con las acciones y con las experiencias.
Pero los periplos neurales no están solo enmarcados por el desarrollo, sino también lo están por la historia más amplia de la evolución. Para comprender este nuevo nivel de encuadramiento, tenemos que estudiar en primer lugar por qué aprendemos.
§. Instintos básicos
La vida de San Jerónimo, un monje del siglo V, se ha elegido a menudo como tema de cuadros religiosos. Los episodios de su vida han sido descritos por artistas como Durero, El Bosco, Leonardo, Caravaggio y Rubens. Pero quizá el retrato más entrañable sea el de Vittore Carpaccio San Jerónimo y el león (figura 79, lámina 11). Según la leyenda, un león llegó cojeando al monasterio de San Jerónimo una tarde, lo que provocó la dispersión de los monjes. Sin embargo, Jerónimo se mantuvo firme y se dio cuenta de que el león sufría porque tenía clavada una espina en una pata. Se la retiró y el león agradecido se convirtió en su amigo, y más tarde le ayudó a proteger a su burro de los peligros.La historia de San Jerónimo es un juego de instintos. Nuestra tendencia natural es huir de un león, aunque Jerónimo supera este instinto al permanecer firme y ayudar al animal. Desde el punto de vista del león, el instinto natural es atacar a los humanos, aunque aprende a reprimir su agresividad y se hace amigo de Jerónimo. Los instintos a menudo se presentan de este modo, como algo negativo que conviene vencer. Pero los instintos en el sentido más amplio se encuentran en el centro de todas nuestras acciones, incluidas las más nobles, porque incluso cuando pensamos que hemos aprendido a vencerlos, dicho aprendizaje está cimentado en nuestras reacciones instintivas. Para ver el porqué, debemos mirar a los orígenes evolutivos del aprendizaje.
La evolución darwiniana se basa en el éxito reproductor. Si las respuestas particulares a los cambios del entorno conducen a un incremento de la supervivencia y de la reproducción, estas respuestas tenderán a diseminarse por la población gracias a la selección natural. Esto explica la prevalencia de las respuestas instintivas tales como la natación de la bacteria Escherichia coli hacia el azúcar, la floración de las plantas en determinados momentos del año, o la respuesta retráctil de las babosas al tocarlas. Todas estas respuestas incrementan el éxito reproductor porque han sido probadas y comprobadas durante muchas generaciones. Pero ¿cuál es la ventaja evolutiva de aprender a responder a retos que quizá nunca antes habíamos encontrado? ¿Por qué la selección natural favorece que un perro aprenda a responder al sonido de una campana, o que un mono responda al sonido de la apertura de una puerta?

Figura 79. San Jerónimo y el león en el monasterio, Vittore Carpaccio, 1501-1509. Véase la lámina 11.
Para que el aprendizaje durante la vida resulte útil para la evolución darwiniana, tiene que establecer predicciones más indirectas sobre el éxito reproductor, algo que consigue gracias a que está ligado a respuestas instintivas. Supongamos, por ejemplo, que tenemos un temor innato a los animales grandes de dientes afilados, una respuesta instintiva que podría haberse reforzado durante muchas generaciones por selección natural. Cuando vemos que un león enseña sus dientes, instintivamente sentimos miedo y echamos a correr. Si ahora aprendiéramos los factores que nos ayudarán a predecir si un león está a punto de enseñarnos los dientes, tal como ver una melena desgreñada o escuchar un gruñido fuerte, podríamos huir sin esperar a ver lo que hace el león, y esa acción incrementará nuestra supervivencia. De igual forma, nuestras posibilidades de supervivencia se incrementarían si pudiéramos aprender que los leones abundan en determinadas áreas, lo que nos permitiría evitarlas o ser cautelosos cuando entremos en ellas.
Tal aprendizaje va a nuestro favor porque el temor instintivo ha sido implantado por la selección natural. Si naciéramos con miedo a las manzanas, aprenderíamos a asustarnos de las peculiaridades que predicen su aparición, como la visión de los manzanos. Pero esto no nos ayudaría a sobrevivir ni a reproducirnos, porque tener miedo de las manzanas no aporta ninguna ventaja selectiva. El aprendizaje aporta ventajas evolutivas cuando está conectado a instintos que la selección natural ha puesto a punto con anterioridad.
Esta relación entre el aprendizaje y el instinto resulta bastante sutil. Los macacos rhesus no nacen con miedo a las serpientes, pero pronto aprenden a tenerlo en cuanto ven que su madre les tiene miedo. Esto parecería sugerir que el miedo a las serpientes no es instintivo, sino que se aprende. Los psicólogos Michael Cook de la Universidad de Wisconsin y Susan Mineka de la Universidad del Noroeste lo exploraron un poco más al mostrar a unos monos lactantes fragmentos de películas de adultos atemorizados y a continuación fragmentos de películas con diferentes objetos[114]. Algunos de estos objetos eran inofensivos, como las flores o un conejo de juguete, mientras que otros eran más cercanos a lo que les resulta potencialmente mortal en la naturaleza, como serpientes o cocodrilos de juguete. Cuando se mostraron estas películas a los monos con menos de dos años, adquirieron un temor a los objetos potencialmente peligrosos, como las serpientes, pero no a las flores ni a los conejos. Evidentemente, estos monos jóvenes ya tienen un marco neural que los predispone a tener miedo de los animales que resultaron peligrosos en el pasado evolutivo de la especie.
Aunque haya ofrecido ejemplos de peligros ambientales que más vale evitar, lo mismo valdrá para los aspectos deseables de nuestro entorno. Instintivamente, preferimos el sabor de determinados tipos de comida, como las manzanas. Si aprendemos que otras peculiaridades, como el color o el olor, son buenos predictores del sabor de las manzanas, entonces podremos utilizar esta información para mejorar nuestra capacidad para conseguirlos. De igual forma, podríamos sentirnos instintivamente atraídos por determinados rasgos del sexo opuesto, como la forma del cuerpo. Si somos capaces de aprender otros rasgos que ayudan a predecir tales formas, como el tipo de ropas que visten o el modo de caminar, entonces nos podrían ayudar a encontrar una posible pareja.
Ayudar a los otros también es un instinto valioso con una base evolutiva. La acción de ayudar a la descendencia o a los familiares podría verse favorecida por la selección natural porque incrementaría el éxito reproductor de los genes compartidos. Ayudar a otros también puede comportar un beneficio cuando se devuelve el favor. En la leyenda, San Jerónimo se beneficia de la ayuda al león porque obtiene un amigo de confianza que le ayuda a proteger su burro. Esto no significa que Jerónimo estuviera pensando en su propio beneficio cuando ayudó al león, sino que el resultado global de ayudar a otro puede resultar ventajoso. Desde un punto de vista evolutivo, el deseo de ayudar a los demás puede plantearse como un instinto más que a veces se verá favorecido por la selección natural. Esto significa que aprender a reconocer a la familia o a los que nos ayudaron en el pasado nos aportará ciertas ventajas.
Todos estos ejemplos se pueden resumir diciendo que nacemos con un conjunto de valores, experiencias que consideramos premios o castigos y que, por lo tanto, desencadenan acciones de compromiso o evasión, valores innatos que nos implantó la selección natural. Los valores iniciales forman parte del patrón de conexiones neurales con las que nacemos y surgen a través del desarrollo. Así pues, el aprendizaje permite que estas conexiones se modifiquen con la experiencia, por lo que incrementamos nuestras posibilidades para obtener lo deseable y evitar lo indeseable.
Nuestros valores iniciales son solo el comienzo de la historia. Tal y como hemos visto, procesos como el aprendizaje DT hacen cambiar las expectativas e introducen nuevos valores (capítulo 7). Para los humanos, el dinero pronostica la capacidad para obtener comida y otros objetos deseables y, por lo tanto, se vuelve valioso por derecho propio. Simbad estaba exultante cuando encontró los diamantes durante uno de sus viajes debido al valor que les atribuye. Los valores también podrían competir entre sí y cambiar según las condiciones. Supongamos que estamos indecisos entre comernos las manzanas de un árbol y huir porque merodea una serpiente. Nuestra decisión estará influida por factores tales como la cantidad de comida que hemos ingerido recientemente. Si tenemos mucha hambre, el valor relativo de las manzanas sube de manera impresionante y es más probable que nos quedemos. Los valores también pueden cambiar con el desarrollo: el interés por las parejas sexuales se incrementa durante la pubertad debido a los cambios hormonales del cuerpo. Existe una interacción compleja entre las condiciones, el desarrollo y el aprendizaje que hace que cambien los valores en todos los sentidos durante nuestros periplos neurales.
No obstante, todos estos cambios se fundamentan en ciertos valores iniciales, unos cimientos que se han ido perfeccionando gracias a la selección natural, porque dichos cimientos proporcionan las razones evolutivas para nuestra capacidad de aprender. Las posibilidades de supervivencia y reproducción se incrementan mediante el aprendizaje para satisfacer los valores instintivos, como la conveniencia de comer cuando se tiene hambre y la inconveniencia del dolor. Si un animal hubiese nacido con el conjunto de valores contrario, deseando el dolor y evitando la comida cuando tenga hambre, el aprendizaje resultaría desventajoso porque el animal aprendería con más eficacia a sortear la comida y a exponerse por sí solo al dolor, lo que rápidamente lo desnutriría o haría que se desangrara hasta la muerte. La selección actuaría en contra de los animales con este comportamiento. Los valores con los que hemos nacido los ha implantado la selección natural y proporcionan los fundamentos del aprendizaje.
§. Flexibilidad frente a rigidez
Hemos visto que el aprendizaje puede ser ventajoso desde un punto de vista evolutivo, aunque también tenga algunos inconvenientes. Primero, el aprendizaje está ligado solo indirectamente al éxito reproductor. Esto significa que podemos comportarnos de forma que no se incremente el éxito reproductor, como decidir no tener niños porque cuesta demasiado. Segundo, el aprendizaje es un proceso mucho más elaborado que las respuestas instintivas, y requiere mayor complejidad de las rutas e interacciones neurales. ¿Por qué evoluciona el aprendizaje cuando ya existen respuestas instintivas más simples y directas?La respuesta está relacionada con el compromiso entre la flexibilidad y la rigidez. Una de las limitaciones de la evolución por selección natural es que solo puede capturar las tendencias del entorno que persisten durante varias generaciones. Pero muchas peculiaridades ambientales cambian en mucho menos tiempo, a menudo de una manera difícil de predecir. Si todas nuestras acciones fueran completamente innatas y no tuvieran en cuenta esta variación del entorno, entonces resultaría difícil alcanzar muchos comportamientos adaptativos. Imaginemos, por ejemplo, que la Venus atrapamoscas tuviese que predecir, sin información de su entorno, cuándo cerrar su hoja sobre un insecto. Necesitaría un modelo complejo del mundo que le dijera con precisión cuándo va a aterrizar un insecto, una predicción que es casi imposible realizar. En vez de prescribir todas las acciones que probablemente incrementen el éxito reproductor, la selección natural ha favorecido estrategias más indirectas.
Un primer paso hacia la vía indirecta consiste en emplear respuestas instintivas, ya que, desde la perspectiva evolutiva, son menos directas que tener todas las acciones predeterminadas desde el nacimiento. La Venus atrapamoscas ajusta sus acciones de acuerdo con el entorno: los movimientos del insecto estimulan el cierre de las hojas. Las respuestas instintivas como esta proporcionan una forma básica de negociar entre la rigidez y la flexibilidad. Al ser instintivas las respuestas, en vez de las acciones, podemos actuar con más flexibilidad ante los cambios de las condiciones. Esta estrategia subyace a un amplio abanico de acciones, como que la bacteria Escherichia coli produzca enzimas que digieren la lactosa cuando están rodeadas de leche, o que nade hacia regiones ricas en azúcares. Estas acciones concretas no están especificadas de antemano, sino que dependen de lo que la bacteria encuentra en el entorno.
Sin embargo, las reacciones instintivas también tienen sus limitaciones, que resultan particularmente evidentes cuando andan dando vueltas por el mundo macroscópico. Un animal pluricelular recién nacido experimenta toda clase de sucesos complejos provocados por sus propios movimientos. Al girar la cabeza, los estímulos visuales cambian de un modo complicado de acuerdo con lo que está a su alrededor. A medida que comienza a caminar, se encuentra con muchos obstáculos por el camino, algunos de los cuales más le valdrá evitarlos y otros le resultarán más beneficiosos. Y para obtener comida, necesita saber dónde encontrarla y cómo manejarse en su complejo entorno para llegar hasta ella. Cada individuo se topa con su propio conjunto de experiencias, de acuerdo con sus contextos y movimientos particulares. Habérselas con toda esta complejidad solo con reacciones instintivas sería muy difícil: el animal tendría que haber nacido con un conocimiento instintivo de todos los tipos de objetos que se podría encontrar, del aspecto que tendrían desde cada ángulo y de cómo ajustar sus movimientos en cada contexto particular. En vez de prescribir todos estos comportamientos mediante respuestas instintivas, la estrategia alternativa consiste en capturar parte de la complejidad a través del aprendizaje, lo que permite que el animal ajuste sus acciones corporales al entorno con mucha más flexibilidad. El animal no necesita tener un conocimiento innato de cómo responder exactamente a los obstáculos y sucesos, sino que modifica gradualmente sus acciones de acuerdo con lo que tiende a ocasionar resultados deseables o indeseables. Aprende a moverse y a ordenar el mundo de determinada manera, de acuerdo con sus experiencias y valores. Las acciones aprendidas es probable que le ayuden al éxito reproductor porque los valores están de por sí enraizados en respuestas instintivas, perfeccionadas por la selección natural. Aunque sea más indirecto que las respuestas instintivas, el aprendizaje proporciona una mayor flexibilidad para manejar la complejidad y las contingencias de lo que se encontrará el animal[115]]. Tener tal flexibilidad incrementará el éxito reproductor y, por lo tanto, ha sido favorecido por la selección natural. Por esta razón han evolucionado nuestra capacidad de aprender y nuestros patrones de aprendizaje.
Al igual que los cuentos de Simbad, el aprendizaje está enmarcado por dos historias más grandes: el desarrollo y la evolución. Pero también hay otra historia más que contar. Nuestros tres cuentos anidados de evolución, desarrollo y aprendizaje enmarcan otro cuento más. Este trata de cómo los humanos, juntos, han provocado otro tipo de transformación: el cambio cultural.
Capítulo 11
El crisol de la cultura
§. El aprendizLas sociedades de hoy en día son muy diferentes de las de hace 10.000 años, no porque haya diferencias importantes entre las propiedades biológicas (si hubiéramos nacido entonces, probablemente encajaríamos perfectamente bien), sino por los espectaculares cambios en la cultura y en las perspectivas. Si pudiéramos viajar en el tiempo como adultos para encontrarnos con nuestros ancestros que vivieron hace diez mil años, nos hallaríamos ante innumerables discrepancias culturales. Los ancestros se sorprenderían inmediatamente de nuestras extrañas ropas, desde los zapatos hasta la camiseta ajustada al cuerpo. Si intentásemos saludarlos con un apretón de manos, probablemente los alarmaríamos y se preguntarían qué estábamos a punto de hacer. A medida que los fuéramos conociendo mejor, quizá aprendiendo algo de su lenguaje, podríamos intentar transmitirles nuestro estilo de vida. Podríamos hablarles de que vivimos en casas con hojas transparentes de un material que deja pasar la luz, de que tomamos la comida con instrumentos que pinchan, y que no solemos recoger nuestra comida, sino que la intercambiamos por trozos de papel. O podríamos describirles que nos movemos muy rápido en cajas con ruedas o volamos por el aire dentro de máquinas aladas, y que hablamos y enviamos mensajes a personas que están muy lejos aunque no podamos verlas. También podríamos intentar explicar que todos vivimos en una enorme masa esférica en rotación y que las estaciones cambian a medida que la esfera va rodeando al sol. Después de escucharnos con incredulidad, quizá nuestros ancestros decidirían mostrarnos algunas pinturas rupestres. Entonces podríamos intentar explicarles que es posible retratar las cosas de otra manera, con el uso de la perspectiva o del color en pinceladas impresionistas. Si cantan para entretenernos, podríamos desplegar un reproductor de música y observar su reacción de curiosidad al escuchar una canción pop, algo de jazz o una sinfonía.
§. Poblaciones fructíferas
§. Cambio duradero
§. Refuerzo cultural
§. La fuerza de la competencia
§. Esfuerzos cooperativos
§. Una mezcla cultural
§. Impulsados por el pasado
§. Una receta cultural
Todos estos contrastes en la ropa, los modales, el estilo de vida, la economía, el transporte, la comunicación, la ciencia y el arte nos indican que las sociedades humanas se han transformado a lo largo de los siglos. Hay muchas maneras de describir la aparición de estos cambios culturales. Podríamos resaltar la importancia de las luchas por el poder, las fuerzas del comercio, el liderazgo de individuos carismáticos, la importancia de la innovación y de la imitación, y que nos resultan cruciales el lenguaje o los medios de transporte y comunicación. Quiero estudiar este problema desde una perspectiva diferente.
Hemos visto la forma de llegar a algunos principios centrales al alejar nuestro punto de vista y examinar lo que es común en un abanico de transformaciones vivientes (evolución, desarrollo y aprendizaje). Ahora quiero incorporar el cambio cultural al equipo. Ya se había propuesto que algunos de los principios que hemos explicado tienen una importancia fundamental en las transformaciones culturales. Los historiadores J. R. y William McNeill, por ejemplo, han insistido en que la competencia y la cooperación fueron decisivas para impulsar el cambio cultural de los humanos[116]. Sin embargo, hasta ahora no se había utilizado la colección completa de ingredientes e interacciones de la receta creativa para la vida para explicar el cambio cultural. Esto se debe a que hay tantos elementos del comportamiento humano entre los que escoger que no es fácil que nos venga a la cabeza el conjunto de los siete principios y sus interacciones. Este cambio cultural solo comienza a cristalizarse cuando optamos por una perspectiva más amplia que abarque todas las transformaciones que afectan a los seres vivos. En tal caso, creo que no solo ganamos un conocimiento más fundamental de la forma en que se introdujo el cambio cultural, sino también una perspectiva más clara de cómo se relaciona con nuestro pasado biológico.
Para transmitir este punto de vista, nos ayudará un ejemplo cultural particular. Podríamos elegir muchas cosas, desde la guerra y la moda a la ciencia y el arte. He elegido un episodio de la vida de Leonardo da Vinci porque quiero demostrar que incluso un ejemplo tan representativo de logro cultural se puede ver a través de los principios que venimos estudiando.
§. El aprendiz
A mediados del siglo XV existía una competencia terrible entre los orfebres Andrea Verrocchio y Antonio Pollaiuolo en Florencia[117] , que por entonces era una ciudad próspera, famosa por sus tejidos y morada de los Medici, la poderosa familia de banqueros. Por lo tanto, los ciudadanos prósperos solicitaban constantemente a los orfebres que fabricaran ornamentos y joyas. Verrocchio y Pollaiuolo compitieron por los mejores encargos en este contexto tan lucrativo. Pero no les bastó con ser los mejores orfebres, sino que ambos querían incrementar sus ingresos y su reputación abarcando nuevas ramas artísticas. Verrocchio decidió captar lo relacionado con las esculturas, mientras que Pollaiuolo se formó como pintor. La primera pintura importante de este último fue una serie de tres grandes lienzos que describen los Trabajos de Hércules, completada hacia 1460 para el palacio de los Medici. Fue una obra a gran escala, que requirió la ayuda del hermano más joven de Antonio, Piero Pollaiuolo. Aunque los lienzos originales se han perdido, una pequeña versión de Hércules y la Hidra, de Antonio Pollaiuolo (figura 80, lámina 12), nos da una idea de su aspecto. Los cuadros de los hermanos Pollaiuolo se consideraron un gran éxito y les reportaron más encargos lucrativos, convirtiéndose en pintores florentinos de renombre.Andrea Verrocchio también tuvo éxito en su nueva profesión como escultor, pero, quizá estimulado por el éxito de su rival, también decidió dedicarse a la pintura al final de la década de 1460. Al no tener un hermano que lo ayudase, reclutó para ello a algunos asistentes y aprendices. Con los años, entre los asistentes formados se encontraban pintores como Sandro Botticelli, Domenico Ghirlandaio y Pietro Perugino, que se convertirían en artistas famosos por propio mérito. Dos de los aprendices eran Lorenzo di Credi y, el más famoso, Leonardo da Vinci. Leonardo nació en 1452 en la ciudad de Vinci en las montañas, a 32 km de Florencia, hijo ilegítimo del jurista Ser Piero da Vinci y una campesina llamada Caterina, y se educó en la casa de su abuelo paterno. Leonardo tenía talento para dibujar desde su más tierna infancia, por lo que su padre decidió colocarlo como aprendiz con Verrocchio cuando Leonardo todavía era adolescente, más o menos cuando Verrocchio se inició en la pintura.

Figura 80. Hércules y la Hidra, Antonio Pollaiuolo, hacia 1475. Véase la lámina 12.

Figura 81. Izquierda: Tobías y el ángel, Andrea del Verrocchio y Leonardo da Vinci, 1470-1480.Derecha: Tobías y el ángel, Antonio Pollaiuolo y Piero Pollaiuolo, 1460. Véase la lámina 13.
§. Poblaciones fructíferas
Leonardo y Verrocchio fueron capaces de hacer lo que hicieron gracias a que pertenecían a un contexto social particular, a una población de individuos. En la época de Leonardo, en Florencia vivían unas 50.000 personas. Esta población y sus ciudades cercanas produjeron algunos de los grandes artistas del Renacimiento, entre ellos, Leonardo, Miguel Ángel y Botticelli[118]. Es poco probable que por entonces circulasen por Florencia un conjunto especial de genes «artísticos», sino que más bien la población florentina del siglo XV representaba una clase social particular que permitió el florecimiento del talento de las personas de diferentes maneras. Quizá hay miles de personas vivas hoy en día con un potencial genético equivalente al de Leonardo o al de Miguel Ángel (la población mundial actual es aproximadamente cien mil veces mayor que la de Florencia en el siglo XV). Estos equivalentes modernos no producen obras maestras como La Gioconda o La creación de Adán porque nacieron y se educaron en un contexto muy diferente[119]. Tal y como hemos visto antes, cada población adopta un contexto determinado, ya sea el juego de la ruleta, un bosque de manzanos, una taza de moléculas de té o el cerebro de una persona. En la historia de Leonardo, el contexto era Florencia y su entorno el siglo XV.La población florentina no estaba formada por individuos idénticos, sino que todos eran diferentes debido a la variación de nacimiento y las circunstancias. Leonardo, Miguel Ángel y Botticelli tenían cada uno unas características genéticas distintas, por lo que cada uno comenzó su vida desde una posición diferente en el espacio neural. Los periplos neurales empezaron también a depender de cada encuentro y cada experiencia acaecidos durante su infancia y después. Su interacción con los familiares, con los compañeros artistas, y con otros ciudadanos, influyeron en los intereses particulares y en las aspiraciones de cada persona. Estas experiencias e interacciones incluyeron sucesos fortuitos, como que Leonardo pidiera trabajo en el estudio de Verrocchio cuando este acababa de iniciarse en la pintura. Como escribió el historiador David Brown: «si Verrocchio no se hubiese puesto a pintar en esa época, seguramente Leonardo nunca hubiera llegado a ser pintor». La población florentina era muy diversa gracias a las complejas maneras en que personas genéticamente diferentes interaccionaban entre sí y con su entorno.
La diversidad humana da opción a muchas posibilidades. Si todo el mundo fuese genéticamente idéntico y se expusiera a exactamente las mismas experiencias, entonces los intercambios entre los individuos serían mucho menos interesantes y fructíferos. Pero como todo el mundo es diferente, podemos aprender y beneficiarnos continuamente del conocimiento y capacidades de los otros. Estos intercambios también influyen en las actividades creativas de cada persona. Como vimos en el capítulo 9, la creatividad se incorpora en el modo en el que los humanos aprendemos del mundo e interaccionamos con él. Los intercambios entre las personas suelen hacer surgir constantemente un amplio abanico de actos creativos. Los logros creativos excepcionales, tal como un cuadro o una idea particulares, comienzan a sobresalir entre la población y se vuelven más evidentes que los demás. Esto permitiría identificar a los individuos que fueran muy creativos. Pero tales individuos nunca operan completamente aislados, pues los avances culturales dependen de poblaciones de personas heterogéneas y creativas que interaccionan entre sí. Al igual que ocurre con las otras transformaciones, el principio de la variabilidad de la población proporciona un ingrediente crítico para el intercambio cultural.
He ofrecido la población de Florencia a modo de ejemplo, pero resulta obvio que esta ciudad no estaba aislada: sus habitantes seguro que viajaban. Leonardo pasó gran parte del final de su vida en Milán y Miguel Ángel viajó a Roma para pintar la Capilla Sixtina. También era posible viajar muy lejos, aunque se tardaran muchos meses. El explorador Américo Vespucio, que nació en Florencia dos años después que Leonardo, hizo varias expediciones a América, que recibió este nombre en su honor. El mundo estaba formado por muchos mundos interconectados y no por poblaciones aisladas[120]. La variabilidad de la población también resulta aplicable a este contexto más amplio en el que se incluyen las diferentes poblaciones mundiales en interacción. Mientras se estaban produciendo algunos avances en Florencia, otros podrían estar ocurriendo en Samarcanda o Pekín. Gracias a la comunicación mundial, estas diferentes poblaciones también pudieron interaccionar e influirse mutuamente. Las pinturas al óleo que usaba Leonardo habían sido inventadas pocos años antes por el artista flamenco Jan van Eyck, y el papel en el que escribía Leonardo era una invención que se remonta a China muchos siglos antes. La población florentina no estaba aislada, sino que era parte de una población más amplia que abarcaba todo el mundo, unos 350 millones de personas en el siglo XV. El Renacimiento en Florencia no habría sido posible sin este enorme telón de fondo de la variabilidad.
§. Cambio duradero
No obstante, por sí solas las variaciones no son suficientes; también se necesita que persistan. El contacto personal y la tradición contribuyen a la persistencia cultural. Verrocchio pudo transmitir su conocimiento al joven Leonardo a través de sus conversaciones y de las enseñanzas prácticas, con lo que le trasladó al pupilo lo que llevaba aprendido en sus años de experiencia. De igual forma, el matemático Isaac Barrow inspiró a su estudiante, Isaac Newton, y el botánico John Henslow enseñó al joven Charles Darwin. Cada pupilo puede a su vez transferir lo que ha aprendido, con lo que el conocimiento y las tradiciones se transmitirán continuamente de generación en generación. El aprender de los demás no se limita, por supuesto, a las clases particulares típicas, sino que ocurre cada vez que interaccionamos y conversamos unos con otros.En la persistencia cultural también pueden intervenir los artefactos. Independientemente de lo original que fuera Leonardo, hubiera tenido poco impacto si sus pinturas, dibujos y escritos se hubieran evaporado en cuanto los hubiera acabado. La producción de utensilios estables desde el punto de vista material proporciona un medio muy eficaz para la transmisión a largo plazo del conocimiento a los otros. Verrocchio y Leonardo pudieron inspeccionar y aprender del Tobías y el ángel de los Pollaiuolo porque los cuadros eran duraderos. Por el mismo motivo, las obras del Renacimiento seguirán inspirando a los artistas en los siglos venideros. Joshua Reynolds, que viajó a Italia para ver tales obras, recomendó en 1769 a los que aspiraban a ser artistas: «reflexiona sobre la manera en que Miguel Ángel o Rafael habrían tratado este tema y actúa con el convencimiento de que ambos artistas verán y criticarán tu cuadro en cuanto lo completes»[121]. De igual forma, Paul Cézanne se inspiró en los viejos maestros con sus regulares visitas vespertinas al Museo del Louvre para observar y bosquejar sus obras. Si retrocedemos aún más, seremos capaces de apreciar el arte rupestre que las sociedades magdalenienses realizaron hace más de 10.000 años (figura 2). Todo esto ocurre gracias a la persistencia del material. Lo mismo ocurre con otros utensilios como la ropa, la escritura o la escultura. Como los productos perduran en el tiempo, las personas pueden aprender de sus predecesores y aprovecharse de sus avances.
La persistencia también se puede potenciar con copias. Hasta la época de Leonardo, la mayoría de las copias se seguían haciendo a mano o con bloques de madera. Pero hacia 1430, después de que Johannes Gutenberg inventara la prensa de impresión de tipos móviles, las copias se fabricaban con mucha más eficacia. Hacia 1500, las prensas de Gutenberg ya estaban en 236 ciudades de Europa y habían impreso unos 20 millones de libros en diferentes idiomas[122]. Leonardo se benefició de esta bonanza impresora: adquirió más de cien libros impresos sobre temas que iban desde las matemáticas, filosofía e historia natural, hasta la poesía. Por lo tanto, la impresión tuvo una importancia decisiva al permitir que las ideas persistieran y se diseminaran. Se siguen desarrollando nuevos métodos de copia, como la fotografía y la distribución electrónica por Internet. Nosotros también podemos generar copias por repetición que se sumarían a las formas de propagación anteriores. La producción en masa de ropa, cubiertos y coches depende de las máquinas que llevan a cabo acciones particulares de forma repetitiva. A diferencia de la imprenta, un coche no actúa como modelo directo de otro, sino que surgen de la repetición continua del mismo proceso de fabricación.
Todas estas formas de persistencia (instrucción personal, utensilios resistentes y copiados) consiguen que los logros culturales tengan efectos duraderos. Sin la persistencia, cada avance individual, independientemente de lo maravilloso que sea, se iría a la tumba con su inventor y tendría poca repercusión en la cultura.
La persistencia cultural y la variabilidad están muy conectadas: ambas giran alrededor de la capacidad de comunicación de los humanos, sobre todo gracias al lenguaje. La comunicación es importante para generar variabilidad mediante los intercambios entre las personas, pero también para la retención y el mantenimiento del conocimiento. La variabilidad por sí sola crearía un frenesí de ideas y acciones efímeras, mientras que si solo existiera la persistencia, conduciría al estancamiento. La cultura consigue cambiarse con eficacia gracias al equilibrio entre la variabilidad y la persistencia.
§. Refuerzo cultural
La variación de la población y la persistencia proporcionan los ingredientes clave para el cambio cultural, pero el impulso para que haya cambios dentro de una población procede de otro ingrediente: el refuerzo. A medida que sus cuadros se exponían en Florencia, la reputación de artistas como los Pollaiuolo, Verrocchio o Leonardo se diseminaba por la población. Si cada persona a la que le gusta un cuadro se lo dice a otras, el mensaje se multiplica como una infección, y a medida que se expande su reputación, los artistas consiguen más encargos lucrativos que les acaban reforzando la fama. El éxito engendra éxito. Incluso los logros de Vincent van Gogh o Paul Cézanne, que no tuvieron mucho éxito durante su vida, se vieron reforzados más tarde por quienes reconocieron y defendieron su obra.Algunos logros se consiguen reforzar en las poblaciones humanas porque las personas tenemos valores comunes: nos suele gustar y disgustar lo mismo. Tal y como vimos en los capítulos anteriores, lo que aprendemos y lo que hacemos depende de lo que valoramos. Podemos valorar muchas cosas (la comida, el sexo, la comodidad, los bienes, los terrenos, el dinero, la categoría social, el conocimiento y la belleza), y como la mayor parte de la población comparte estos valores, los logros que los satisfacen tienden a reforzarse y a diseminarse.
Sin embargo, no todos los individuos de una población comparten exactamente los mismos valores debido a las variaciones de nacimiento y a la educación. Algunas personas darán más valor que otras a determinadas cosas, como a los bienes, al dinero o al conocimiento. Los especialistas también valorarán cosas que para otros son menos evidentes: cuando miran los cuadros de los hermanos Pollaiuolo, de Leonardo y de Verrocchio, sin ninguna duda verán y valorarán particularidades que la mayoría no apreciaremos. Las poblaciones humanas poseen una colección de valores heterogéneos. Sin embargo, dado que muchos son comunes en determinados grupos sociales, los logros que satisfacen esos valores se diseminan a través del refuerzo, al menos dentro de estos grupos.
Lo que valoramos también depende enormemente del contexto cultural. Cuando Leonardo tenía 30 años se mudó a la ciudad de Milán, gobernada por la familia Sforza. A diferencia de los Medici de Florencia que eran banqueros, los Sforza eran militares que obtuvieron su poder gracias a acciones de armas. Cuando Leonardo escribió su carta de presentación al duque de Milán, Ludovico Sforza, la elaboró con los intereses militares del duque en mente:
Mi Ilustrísimo Señor: habiendo visto y examinado cumplidamente las invenciones de todos los que se consideran a sí mismos fabricantes y maestros de dispositivos de guerra, he encontrado que el diseño y el funcionamiento de sus máquinas no son en absoluto diferentes de lo que se usa habitualmente. Por lo tanto, y sin querer ofender a nadie, me atrevo a ofrecerle mis conocimientos a Su Excelencia y a hacerle partícipe de mis secretos, y estaré encantado de demostrarle su eficacia en el momento en el que estime conveniente.[123]A continuación, Leonardo pasó a enumerar las invenciones que había ideado que pondrían a los Sforza a la cabeza de las tácticas militares. Entre ellas se encontraban puentes militares portátiles, métodos para extraer agua de las trincheras durante los asedios, métodos para destruir fortalezas, nuevos tipos de cañones y diseños para carros armados. La lista mostraba lo que Leonardo pensó que Ludovico Sforza encontraría particularmente valioso. Los intereses de los Sforza a su vez se reflejaban en un conflicto militar que prevalecía entre familias y estados rivales de la Italia en la época. El propio Leonardo fue testigo de la expulsión militar de los Sforza por los franceses varios años después de haberse mudado a Milán.
Se trata de un ejemplo que muestra que los valores culturales no están fijos, sino que dependen del contexto, de las agrupaciones sociales y de los problemas dominantes en la época. Sin embargo, dado que los pueden compartir muchos individuos de una población o grupo social, conducen al refuerzo de ideas y logros que satisfacen dichos valores comunes. El refuerzo es un ingrediente clave del cambio cultural porque, gracias a él, se favorece el ascenso de determinados logros en una población.
§. La fuerza de la competencia
El éxito a menudo conduce a la competencia: a medida que se difunde una idea o logro y la van conociendo todos, la novedad se disipa. El logro comienza esencialmente a competir contra sí mismo porque a medida que la mayoría de una población ya está familiarizada con él, disminuye la velocidad a la que se disemina. Los logros exitosos también estimulan la competencia de los otros. Cuando Verrocchio fue consciente de lo bien que lo estaban haciendo los hermanos Pollaiuolo, se dedicó a la pintura para poder también llevarse una parte de ese lucrativo mercado. Es más, el éxito de los Pollaiuolo ayudó a la competencia, pues al observar detenidamente Tobías y el ángel de los Pollaiuolo, Verrocchio y Leonardo se beneficiaron de sus avances y pintaron una versión mejorada. Para los Pollaiuolo, que sus logros llamaran la atención de otros fue un arma de doble filo: la atención era una medida de su éxito, pero también provocó y promovió la competencia. De igual forma, el éxito y la fama que Leonardo conseguiría más tarde estimuló los logros de sus competidores, como Miguel Ángel[124]. Se trata de otro ejemplo de nuestro doble bucle de realimentación, en el que el refuerzo mejora la competencia (figura 82). Un logro valioso se disemina mediante la comunicación, pero este éxito es auto limitante porque la novedad desaparece a medida que la población comparte el conocimiento, y se estimula la competencia. La competencia cultural surge por las distintas limitaciones: hay límites para el tamaño de las poblaciones humanas, para el número de logros que estas valoran mucho, y para las recompensas que de ellos se pueden obtener. La sociedad florentina reconoció que eran realmente destacables solo unos pocos artistas, y las familias ricas, como los Medici, encargaron un número limitado de obras, limitaciones que inevitablemente provocaron la competencia. Si la población fuese infinita, entonces la novedad de un logro nunca desaparecería porque podría diseminarse continuamente a nuevos oídos. Y si los encargos fueran ilimitados y el reconocimiento inagotable, entonces apenas habría motivos para que compitieran Verrocchio y los Pollaiuolo.Incluso cuando pensamos que un individuo está trabajando movido simplemente por la curiosidad o por la búsqueda de satisfacción personal, la competencia se deja ver cuándo se encuentra en peligro el éxito o el reconocimiento. Darwin se pasó muchos años desarrollando su teoría de la evolución por medio de la selección natural sin publicar sus ideas. Pero se alarmó cuando recibió una carta de Alfred Russel Wallace en la que le presentaba la misma teoría, y temió que Wallace pudiera llevarse todo el crédito. Darwin encontró una solución amistosa y publicaron sus hallazgos juntos, pero la historia resalta que la competencia puede acechar detrás de cualquier escenario. De igual forma, cuando Leonardo mencionó sus conocimientos secretos sobre máquinas de guerra en su carta al duque de Milán, era claramente consciente de que los competidores podrían robarle sus ideas y, por lo tanto, privarles a él y al duque de su ventaja.
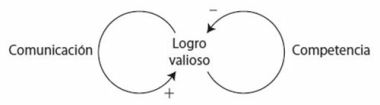
Figura 82. Interacción entre el refuerzo (bucle positivo) y la competición (bucle negativo) en la cultura humana.
§. Esfuerzos cooperativos
Que Leonardo entrara a trabajar en el taller de Verrocchio resultó beneficioso para ambas partes. Leonardo adquirió los importantes conocimientos de su profesor, mientras que Verrocchio se aprovechó de la ayuda de su aprendiz. El cuadro Tobías y el ángel, en el que ambos artistas pintaron su parte, constituye un claro exponente de esta cooperación. Que se encontraran en el mismo lugar fue algo determinante para su relación. Como con otros casos de cooperación, la proximidad física proporciona condiciones que alientan la ayuda mutua y a compartir los beneficios. La cooperación desempeña una función clave en el cambio cultural a muchos niveles. Nuestra capacidad para hablar y comunicarnos procede de la cooperación entre los humanos. Si nos educaran completamente aislados, entonces no aprenderíamos ningún lenguaje elaborado ni tendríamos la capacidad de comunicarnos con eficacia con los demás. Sin la capacidad para comunicarnos, seríamos incapaces de sacar ningún beneficio de los otros, además de que sería poco probable que nuestros logros persistieran y se diseminaran[125]. La existencia del lenguaje y de la comunicación, cuya intervención es crucial para el cambio cultural, se debe a que los humanos hablamos e interaccionamos entre nosotros en grupos. De igual forma, nuestra predisposición a intercambiar dinero y bienes es un riesgo cooperativo[126]. Leonardo se mantenía gracias a los encargos y pagos de familias como los Medici o los Sforza. Y a la inversa, los que adquirieron sus obras se beneficiaron de poseer cuadros prestigiosos de gran calidad. Sin un contexto económico así de intercambio y cooperación, Leonardo no habría sido capaz de mantenerse ni de conseguir todo lo que le ha hecho famoso.También se producen otros niveles de cooperación. Además de formar parte del taller de Verrocchio, Leonardo era miembro del gremio de los artistas florentinos. Se trataba de uno de los muchos gremios de la época, como los vendedores de telas, banqueros y médicos, que velaban por los intereses de sus miembros. Leonardo también era un ciudadano de Florencia, otro grupo definido por intereses comunes. Estos diferentes grupos se desarrollaron mediante la interacción entre la cooperación y la competencia. La cooperación entre Leonardo y Verrocchio estaba promovida por la competencia: Verrocchio necesitaba aprendices para producir obras que rivalizarían con las de los hermanos Pollaiuolo. Y otras agrupaciones, como los gremios o las ciudades, también se mantenían gracias a la rivalidad. Al ser principalmente un pintor, Leonardo no se avergonzaba de promocionar su propio medio en comparación con los otros: «La escultura revela lo que es con poco esfuerzo; la pintura parece algo milagroso que hace que las cosas intangibles parezcan tangibles, las presenta en relieve aunque sean planas, y acerca a la mano lo distante»[127]. Por supuesto, los expertos en otras áreas podrían despreciar la pintura tal y como Leonardo despreció la escultura. El poeta Lord Byron expresó su punto de vista sobre la pintura: «Sin duda, de todas las artes, es la más artificial y antinatural, y aquella por la cual más se impone el sinsentido del género humano»[128]. Tal menosprecio de los grupos competidores no se limita a las artes. El físico experimental Ernest Rutherford, descubridor del núcleo atómico, tendía a minimizar la contribución de los colegas que tenían inclinaciones más teóricas. Se suele citar textualmente que dijo que los teóricos «enredan con sus símbolos, pero los que estamos en el Cavendish sacamos los hechos reales de la naturaleza»[129]. Tal y como hemos visto en los capítulos anteriores, la cooperación puede engendrar competencia y verse mantenida por ella.
Está muy claro que Leonardo apreciaba la interacción entre la cooperación y la competencia. En sus escritos sobre pintura consideró las diferentes ventajas de trabajar junto a otros:
Digo y estoy preparado para demostrar que, cuando dibujamos, es mucho mejor estar en compañía de otros que en soledad, por muchas razones. La primera es que nos avergonzaremos de estar entre los ilustradores si estos nos superan, y esta sensación de vergüenza nos hará progresar en el estudio. En segundo lugar, una envidia sana nos estimulará para que nos unamos a quienes son más elogiados que nosotros, porque sus alabanzas nos servirán de acicate. Además, adquiriremos algunas costumbres de aquellos cuya obra es mejor que la nuestra, mientras que si fuéramos mejores que los demás, nos aprovecharíamos de ver cómo evitar sus errores, y los elogios de los otros tenderán a mejorar nuestra posición.[130]Para Leonardo, que los artistas trabajen juntos dará lugar a una mezcla creativa de interacciones competitivas y cooperativas, y resultará claramente más eficaz que cuando están cada uno por su lado.
La cooperación y la competencia hacen pareja en el cambio cultural. La cooperación permite que los humanos aprendan de los demás y se beneficien de ellos a diferentes niveles, desde la capacidad de comunicación de las personas hasta una mayor cooperación. Si se promocionan los valores y aspiraciones compartidos, se ayuda a reforzar los logros. La competencia desempeña una función clave a la hora de mantener la cooperación, además de incitar continuamente a la consecución de mayores logros.
§. Una mezcla cultural
Al reunir a las personas, la cooperación también alienta la combinación de ideas y habilidades de nuevas maneras. No inventamos desde cero, sino que combinamos y nos fundamentamos en lo que ya conocemos. Cuando Leonardo trabajó en el taller de Verrocchio, siempre integraba sus ideas y observaciones con lo que aprendía de su maestro y de los otros discípulos que trabajaron a su alrededor. Sus logros representaron una combinación original de habilidades y conocimiento, y no algo completamente nuevo. Para Joshua Reynolds, «la invención, estrictamente hablando, es poco más que una nueva combinación de las imágenes que se han reunido y depositado previamente en la memoria»[131]. Este es el principio de la riqueza combinatoria, pero ahora en forma de personas, de sus ideas y de sus logros.La intervención de la riqueza combinatoria resulta más evidente cuando miramos al medio por el que se propagan los logros culturales. El abecedario escrito tan solo consta de 26 letras, pero esto no nos impide expresar ideas nuevas y complicadas en la poesía, en la literatura y en la ciencia. Cuando llegamos a un conocimiento nuevo, no sentimos la necesidad de inventar letras nuevas porque el número de combinaciones posibles con lo que ya tenemos es casi ilimitado. Lo mismo ocurre con las palabras: de vez en cuando se acaban necesitando algunas nuevas, como el término gen, acuñado por Wilhelm Johannsen en 1909. Pero suele bastar con las palabras que ya tenemos: combinándolas de determinadas maneras se expresan y comunican las ideas nuevas. Y lo mismo vale para las notaciones musicales o matemáticas. El dibujo es también una actividad combinatoria: los artistas mezclan unos pocos colores en su paleta y los distribuyen en diferentes combinaciones para conseguir un efecto maravilloso. La riqueza combinatoria es evidente en muchos planos de la cultura, desde la disposición de las letras y de las palabras de un idioma hasta la yuxtaposición de ideas o materiales, o los encuentros entre diferentes personas.
En los capítulos anteriores, el gran número de posibilidades que surge mediante la riqueza combinatoria se transmitía a través de espacios multidimensionales. En la evolución, las poblaciones viajaban por el espacio genético, un embrión en desarrollo vivía una odisea por el espacio del desarrollo, y durante el aprendizaje, el cerebro viajaba por el espacio neural. Ahora estamos en condiciones de pensar que la especie humana viaja a través del espacio cultural, que representa el enorme abanico de posibilidades culturales. En algún momento podemos pensar que la especie humana (todas las personas de la Tierra con sus diferentes intereses y logros) representa una posición en este gran espacio cultural inimaginablemente grande. Con el paso del tiempo, nuestra especie ha viajado por el espacio cultural y ha tomado un camino determinado a medida que las costumbres y esfuerzos humanos se han ido modificando por todo el mundo[132].
Cuando vemos el periplo de nuestra especie por el espacio cultural, resulta tentador verlo como una progresión que nos ha conducido a la cultura actual. Sin embargo, que veamos un cambio como progreso o no depende de nuestros valores concretos, que pueden a su vez cambiar con el tiempo. Además, el periplo de nuestra especie no ha tomado un camino lineal sencillo, sino tortuoso con muchas vueltas y revueltas. Por lo tanto, resulta engañoso pensar en el periplo humano como algo que inevitablemente se veía atraído hacia el presente. Aun así, podemos resumir brevemente algunas de las etapas clave que tuvieron lugar por el camino.
Incluso antes de que nuestra especie Homo sapiens se embarcase en este periplo cultural, las especies relacionadas con la humana se embarcaron en algunos viajes preliminares. Los primeros utensilios humanos, los cantos tallados de África, datan de hace unos dos millones de años[133]. Se cree que están construidos por el Homo habilis, una especie con un cráneo más pequeño (casi la mitad de volumen) que el nuestro. Las herramientas que dejaron detrás sugieren que estos humanos ya eran capaces de pensar en el futuro, ya que cualquier herramienta de piedra está fabricada con un propósito ulterior en mente; o sea, tiene valor a través de la recompensa demorada que ofrecerá: la carne que su usuario podrá cortar de una presa. Este valor lo habrían compartido los individuos de la especie, y suponiendo que también tuviesen cierta capacidad de comunicación, las herramientas y su fabricación se podrían haber extendido por las primeras poblaciones humanas mediante el refuerzo. La necesidad de producir mejores herramientas puede haber conducido también a la competencia, y la cooperación podría verse incrementada cuando un fabricante de herramientas aprende de otro. En los años siguientes, los humanos fueron fabricando mejores herramientas y produjeron un abanico de hachas manuales muy eficaces, en parte debido a los cambios evolutivos de la capacidad encefálica, pero también por las mejoras de la fabricación que se transmitieron por vía cultural. La primera especie humana ya estaba realizando su periplo por el espacio cultural.
Los humanos modernos, la especie Homo sapiens, se cree que han evolucionado hace unos 200.000 años en África, desde donde comenzaron a extenderse como una mala hierba por todo el globo, desplazando a otros parientes, como el Homo neanderthalensis. No está claro por qué la dominación del Homo sapiens comenzó a afirmarse durante este periodo. Sin duda alguna, intervinieron en el desarrollo del lenguaje y de la comunicación, pero todavía no hay acuerdo sobre la manera exacta en la que el Homo sapiens se convirtió en la especie humana dominante[134]. Además, como el cambio cultural conlleva la interacción entre muchos ingredientes, probablemente sea un error basarlo en una única causa. Cualesquiera que sean las razones, hace unos 10.000 años, el Homo sapiens era la única especie humana que quedaba en el planeta. A continuación, nuestra especie colonizó todos los continentes excepto la Antártida, y formó una población mundial de unos pocos millones. Fue aproximadamente entonces cuando los humanos aprendieron a cultivar las plantas y a domesticar los animales. Las semillas y el ganado se volvieron valiosos debido a que eran una promesa de comida para el futuro, y comenzaron a desarrollarse y a diseminarse las habilidades relacionadas con la producción agrícola. El desarrollo de la agricultura representa una etapa clave en el periplo de nuestra especie por el espacio cultural porque proporcionó estabilidad y un excedente de comida que permitió innovar más para desarrollarse y extenderse. Con la mejora del transporte y de las técnicas de cultivo, los humanos comenzaron a formar grupos organizados más grandes, o civilizaciones, hace unos 5.000 años por los valles fértiles de ríos, como el Nilo en Egipto, el Tigris y el Éufrates en Mesopotamia (Iraq moderno), el Indo en Pakistán y el Huang He (río Amarillo) en China. A medida que las civilizaciones cooperaban y competían, y emergían otras nuevas, continuó el periplo por el espacio cultural con la exploración de nuevas direcciones ayudados por innovaciones como la escritura y el dinero. A medida que los humanos iban mejorando su capacidad para cultivar comida y explotar los recursos, también iban incrementando en número: hace 2.000 años, la población mundial había alcanzado aproximadamente los 200 millones. La expansión de la población ha continuado hasta hoy en día, espoleada en el siglo XIX por nuestra mayor capacidad para explotar la energía de los combustibles fósiles, y en el siglo XX por el desarrollo de medicinas, de la higiene y de la agricultura intensiva. Nuestra posición en el espacio cultural actual es producto de las costumbres, aspiraciones y logros de 7.000 millones de personas. Por supuesto, nuestra especie sigue desplazándose por el espacio cultural.
La especie humana ha tomado una senda concreta en un espacio cultural inimaginablemente inmenso. ¿Qué es lo que nos impulsa continuamente?
§. Impulsados por el pasado
Todos los principios que hemos tratado/abarcado hasta ahora en este capítulo funcionan dentro de un contexto social determinado. El tipo de variación de las poblaciones humanas, su persistencia, los valores que la refuerzan, el modo en que las personas compiten, cooperan y combinan sus esfuerzos, todos ellos se aplican a entornos sociales concretos. Uno de ellos fue la población de Florencia en el siglo XV. Pero estos contextos, o entornos sociales, no están fijos, sino que cambian mediante los mismos procesos que ya hemos tratado. Al responder al cuadro de los Pollaiuolo Tobías y el ángel, Verrocchio y Leonardo modificaron el contexto artístico al crear su propia versión: su cuadro proporcionó un nuevo estándar al que tenían que reaccionar los otros. La misma historia se aplicará a otros artistas en el futuro, desde Miguel Ángel hasta Monet. Incluso cuando alguien reacciona negativamente ante sus precursores, como cuando los cubistas se rebelaron contra las normas de la perspectiva, las obras anteriores tienen una importancia fundamental a la hora de engendrar esa reacción. Cada contexto proporciona semillas para el siguiente.Se pueden explicar del mismo modo los logros militares. Dado que los cañones ya se habían inventado y su uso estaba generalizado, Leonardo fue capaz de idear determinadas mejoras que todos los ejércitos comenzaron a utilizar en cuanto las conocieron, con lo que se necesitaron, a su vez, otras mejoras para ser más competitivos. Por este motivo, Leonardo recalcó la confidencialidad en su carta al duque de Milán. Cada nueva invención militar no solo se basó en las anteriores, sino que también impulsó el proceso al cambiar el contexto. De igual forma, en cuanto se establece una teoría científica, proporciona los fundamentos para más investigaciones y desarrollos científicos, lo que llevará a mejorar la teoría o a sustituirla por otra nueva y mejor. Los contextos proporcionan semillas para su propio cambio porque no solo conducen a respuestas concretas, sino que esas mismas respuestas cambian a su vez el contexto.
Los desplazamientos culturales son relacionales, siempre modifican lo que ya existía, y en cada etapa cambian el contexto y los estándares, y por lo tanto, proporcionan el impulso necesario para el siguiente. Esto no ocurre en un único lugar o disciplina, sino que sucede una y otra vez por todo el mundo en muchas áreas de actividad cultural, entre ellas las artes, la tecnología, la ciencia, la moda o el comercio. Es nuestro conocido principio de recurrencia aplicado a las iniciativas humanas.
Estos cambios autopropulsados en el espacio cultural no solo implican la diseminación de los nuevos logros, sino también cambios en algunos de los valores que refuerzan dichos logros. Leonardo tenía varios criterios para juzgar el éxito de la obra de un pintor: «Primero hay que considerar si las figuras tienen el relieve que requiere su posición… Segundo, hay que considerar si la distribución o disposición de las figuras está concebida de acuerdo con las condiciones que se desean representar. Tercero, si las figuras se muestran entregadas a su propósito»[135]. Aunque fueran apropiados para el arte del Renacimiento, con su énfasis por capturar la realidad y la perspectiva, estos valores prácticamente no se aplican a los pintores impresionistas o cubistas que vinieron después. Lo que valoramos, nuestros criterios de éxito, cambian a medida que respondemos a lo que había antes y construimos sobre ello.
Esto no significa que todos los valores estén cambiando continuamente: a lo largo de las diferentes épocas, los pintores compartieron sin ninguna duda algunos valores, como el amor a la pintura y a la observación. Podemos ofrecer una historia coherente de la pintura gracias a tales coincidencias en los puntos básicos. Cada artista reaccionará ante sus colaboradores y predecesores, e introducirá nuevos elementos en el campo. El impresionismo no apareció como caído del cielo, sino que apareció fundamentado en lo que ya había. No obstante, decir que un cuadro de un impresionista como Monet es mejor que el de un artista del Renacimiento, como Leonardo, se consideraría un juicio subjetivo porque no hay un criterio fijo o común que nos permita emitirlo: los valores con los que evaluamos un cuadro han ido variando. Incluso podemos emplear varios valores al mismo tiempo y decidir que tanto Monet como Leonardo son grandes artistas, cada uno en su propio estilo. Las modas en la forma de vestir son otro ejemplo de variación de algunos valores. El valor básico de tener buen aspecto puede no cambiar, pero el tipo concreto de ropa que valoramos variará de un año a otro. Algunos valores son más resbaladizos que otros y se alteran con más rapidez, impulsados por el propio cambio cultural. La ciencia se basa en un conjunto relativamente fijo de valores centrales que se basan en lo bien que las teorías explican las observaciones, o lo simples que son. Estos valores no siempre han estado presentes. Leonardo siempre insistió en la importancia de la observación para formarse opiniones científicas, en vez de basarse en las creencias tradicionales y en los rumores. De hecho, durante el Renacimiento comenzaron a emerger los valores científicos modernos. Una vez que estos valores comenzaron a establecerse y a impregnar con firmeza la cultura, permitieron juzgar los logros científicos mediante un conjunto relativamente fijo de criterios. Somos capaces de explicar el progreso en la ciencia porque nos preguntamos qué teoría proporciona la mejor y más sencilla explicación de lo que observamos, lo que conduce a que la teoría y las observaciones casen mejor. De igual forma, podemos explicar las mejoras de la tecnología, como la comunicación, porque hemos aceptado las medidas a las que nos podemos referir, como la velocidad a la que viaja la información. En la medida en que poseen un amplio conjunto de criterios fijos con los que juzgar las cosas, la ciencia y la tecnología son similares a procesos como la evolución y el aprendizaje predictivo. En la evolución, los organismos o genes se «juzgan» según los criterios de éxito reproductivo, mientras que en el aprendizaje predictivo, los cambios neurales se «juzgan» según los criterios de obtención de recompensas.
Otra peculiaridad de los desplazamientos culturales recurrentes es que se realimentan para alterar la eficacia de los principios en los que se fundamentan. Una de las tendencias principales de nuestro periplo cultural ha sido el incremento de la velocidad y el alcance de las comunicaciones y el transporte. Nuestra capacidad para explotar los caballos y los camellos nos permitió expandir el alcance y la interacción entre las primeras civilizaciones sobre la tierra. De igual forma, las innovaciones en el transporte marítimo y en la navegación fueron clave para permitir el transporte de bienes e ideas entre diversos países. Estos desarrollos están movidos por el valor que los humanos dieron a su tierra y a los bienes. Pero al conectar con más eficacia las diversas poblaciones, estos desarrollos también dieron lugar a que los ingredientes del cambio cultural fueran más eficaces: incrementaron la variabilidad al incrementar el tamaño de la población que es capaz de interaccionar; permitieron que los logros se extendieran cada vez más lejos mediante la persistencia y el refuerzo; incrementaron el margen de competencia y de cooperación; y promocionaron más niveles de riqueza combinatoria al poner en contacto a personas diferentes entre sí. El efecto colectivo fue acelerar nuevos desarrollos técnicos para el transporte y la comunicación, lo que finalmente condujo a trenes, coches, aviones, teléfonos y a Internet. Cada una de estas innovaciones se basó en lo que ya existía: el primer nombre para el automóvil fue coche sin caballos, y una radio se conoció como un inalámbrico, todos ellos nombres extraños a menos que conozcamos su historia. El efecto neto de estas mejoras en las comunicaciones y en el transporte fue que el mundo se encogió con la expansión de la población de humanos, lo que aceleró por realimentación el cambio cultural y a su vez mejoró la eficacia de los principios en los que se basa. El ritmo siempre creciente del cambio cultural del que somos testigos hoy en día es el resultado de tal realimentación.
§. Una receta cultural
Hemos visto que el cambio cultural funciona de acuerdo con la misma receta que nos hemos ido encontrando a lo largo de este libro. La fuerza impulsora para el cambio es el doble bucle de realimentación entre el refuerzo y la competencia, de manera que el éxito se favorece a sí mismo y provoca sus propias limitaciones. Estos bucles están alimentados por un equilibrio entre la variabilidad de la población (que genera continuamente nuevas ideas y yuxtaposiciones) y la persistencia (permite que los logros se mantengan y diseminen a través de la población). La cooperación también resulta esencial al permitir que las personas se beneficien de las habilidades de los demás, lo que favorece los logros dentro de los grupos y también produce nuevos niveles de competencia entre ellos. Al reunir a las personas con las ideas, la cooperación también conduce a un número enorme de posibilidades combinatorias, lo que crea un inmenso espacio cultural a través del cual se desplaza nuestra especie. Este movimiento está auto impulsado, lo que redefine continuamente su propio contexto mediante el principio de recurrencia.Se trata de los mismos principios e interacciones que encontramos en los fundamentos de la evolución, del desarrollo y del aprendizaje. En la evolución, afectan a poblaciones de individuos que se reproducen en un entorno. En el desarrollo, los principios actúan sobre poblaciones de moléculas o de células dentro de un embrión en crecimiento. En el aprendizaje, actúan sobre poblaciones de activaciones neurales en el cerebro. Con el cambio cultural, nuestra receta se aplica a poblaciones de seres humanos en interacción. En cada sistema vemos poblaciones que se impulsan a sí mismas por los inmensos espacios combinatorios: genético, del desarrollo, neural y cultural. El cambio cultural es nuestro cuarto ejemplo de receta creativa para la vida.
Debemos ser cuidadosos y no intentar llevar estas similitudes demasiado lejos. El cambio cultural es diferente en muchos aspectos a los otros procesos: un acto creativo es muy diferente a una mutación o a una colisión molecular, y los patrones de la transmisión cultural debidos al lenguaje, a los utensilios y al copiado no son lo mismo que las leyes de la herencia genética o que la modificación sináptica. Ha habido intentos de conectar con más precisión la cultura y los procesos biológicos relacionados. Por ejemplo, se ha propuesto que una unidad de transmisión denominada meme desempeña en la cultura una función parecida a la del gen en la evolución[136]. Un meme es una idea o actividad que se replica y se transmite de un humano a otro, igual que un gen puede pasar de un padre a su hijo. Mientras que el concepto de meme puede ser útil para resaltar ciertos paralelismos entre la evolución y el cambio cultural, también invita a la confusión. Por ejemplo, ¿cuál es el equivalente preciso de la selección natural que hace que un meme se incremente a expensas de otro? La respuesta dista mucho de estar clara, lo que condujo al genetista Jerry Coyne a concluir que el problema con la memética es que «parece completamente tautológica, incapaz de explicar por qué se extiende un meme, excepto al afirmar, a posteriori, que tenía cualidades que le permitían extenderse»[137].
Es importante detenerse a reflexionar para ver la relación entre la cultura y los otros procesos con un nivel adecuado de abstracción. Gracias a los principios comunes globales de la receta creativa para la vida veremos su similitud en la forma. Pero que seamos conscientes de las similitudes formales no significa que debamos buscar una correspondencia recíproca entre los componentes. Reconoceremos que el ajedrez y la guerra son contiendas territoriales sin que se nos ocurra hacer coincidir los escaques del tablero con las unidades de superficie en una batalla. De hecho, ponernos a buscar la correspondencia entre las unidades de superficie sería un intento descaminado que nos distraería en vez de ayudarnos a comprender estos procesos.
Al resaltar las similitudes en la forma en que se aplican a los diferentes tipos de transformaciones de los seres vivos, he intentado mostrar que la receta creativa para la vida proporciona un nivel útil y apropiado de abstracción. Así pues, ya somos capaces de ver que hay un conjunto común de principios e interacciones en el centro de cada proceso. También sabemos apreciar que, como humanos, hemos tenido el privilegio de formar parte de cuatro periplos extraordinarios: el periplo por el espacio genético que condujo a la especie humana; el periplo por el espacio del desarrollo que condujo a nuestro nacimiento y crecimiento; el periplo por el espacio neural que condujo a nuestros pensamientos y acciones; y el periplo por el espacio cultural que nos permite disfrutar de las ideas y de los logros de los demás. Somos tanto pasajeros como participantes de estos cuatro grandes viajes de la vida. Además de tener una forma parecida, estos periplos también están conectados de otra manera. Los ingredientes básicos del cambio cultural surgieron de los primeros procesos de la evolución, del desarrollo y del aprendizaje: nuestro periplo cultural sigue a los otros tres. Pero también hay una relación que va en otra dirección: nuestra comprensión científica de la evolución, del desarrollo y del aprendizaje en sí misma es un producto cultural, y gracias a la cultura vemos todas las transformaciones de los seres vivos. Para conocer mejor esta relación recíproca entre la cultura y las otras transformaciones, necesitamos mirar más de cerca cómo están conectados los cuatro periplos.Contenido:
§. Orígenes culturalesLos humanos somos magníficos contadores de cuentos. Gracias a las narraciones solemos comunicar nuestra sabiduría y nuestros conocimientos. Nos gusta que nuestras narraciones tengan diferentes estructuras, con comienzos y finales claros, y nos suele gustar que las cosas ocurran por razones concretas que podamos disponer en una secuencia. Estas consideraciones también serían válidas para una historia sobre nosotros mismos. Si seguimos la historia de la humanidad, nos solemos preguntar de forma natural cuándo comenzó y qué nos hace únicos. A menudo buscamos respuestas que nos definan cualidades como el lenguaje, la imitación, la capacidad para planificar, el alma humana o la creatividad. Pero a lo largo de este libro, cuando hemos estudiado el origen de las transformaciones de lo viviente, no hemos encontrado puntos de partida claros ni esencias que lo definan. No hay ningún inicio obvio ni cualidades únicas que expliquen la evolución ni el desarrollo ni el aprendizaje, sino que surgen mediante una colección de ingredientes que se reúnen e interaccionan entre sí. En vez de puntos de partida claros, hemos encontrado bucles e interacciones.
§. Mundos posibles
§. Autorretrato de la naturaleza
Lo mismo le ocurre al cambio cultural: nuestro panorama actual no se puede rastrear para llegar a una causa última, sino que depende de una receta con muchos ingredientes en constante interacción. En vez de una fecha concreta de inicio o de una esencia que lo defina, el origen de la cultura humana se encuentra en la manera en que surgen y se reúnen sus distintos componentes. Echemos un vistazo a la procedencia de cada uno de los principios que hay detrás de las transformaciones culturales.
§. Orígenes culturales
La variabilidad de los humanos surge de las diferencias al nacer y al educarnos. El cerebro con el que nacemos está influido por los genes que heredamos y por la manera en que modifican nuestro desarrollo en el útero. Después de nacer se adquieren más características individuales a medida que nos encontramos en distintas situaciones y aprendemos de ellas. Mediante estos encuentros, nuestro cerebro se va construyendo sobre expectativas y discrepancias, lo que nos convierte en individuos con una determinada manera de ver y hacer las cosas. La variabilidad que permite el cambio cultural en las poblaciones humanas se puede seguir hasta la naturaleza del cerebro y de sus interacciones, lo que a su vez es reflejo de la interacción entre la evolución, el desarrollo y el aprendizaje.Lo mismo vale para la persistencia cultural. La transmisión del conocimiento y la experiencia de una persona a otra depende de nuestra capacidad para comunicarnos. Esta habilidad, a su vez, se fundamenta en el desarrollo y el aprendizaje de cada uno de nosotros. El cerebro nos permite aprender idiomas, ademanes y acciones, y de esta forma establecer una comunicación eficaz con los demás. También podemos aprender a fabricar herramientas u otros utensilios duraderos, y ayudar a transmitir el conocimiento entre las personas. La persistencia cultural está anclada en el modo en el que nuestro cerebro evolucionó, se desarrolla y aprende, lo que nos permite comunicarnos satisfactoriamente y con consecuencias duraderas.
Nuestra capacidad de comunicación también desempeña una función esencial en el refuerzo cultural. Gracias a ella, determinadas ideas y logros se diseminan por una población. Pero lo que se disemina también depende de lo que valoramos: nacemos con unos valores determinados, impregnados en las conexiones neurales de nuestro cerebro, que están enraizados en nuestro pasado evolutivo. Nuestros deseos básicos de comodidad, de comida y de pareja surgen porque tienden a promover la supervivencia y la reproducción. Gracias al aprendizaje, estos y otros valores innatos se elaboran y amplifican de distintas maneras. Podemos llegar a valorar el dinero, la fama, el conocimiento o un cuadro bonito, pero cualesquiera que sean los valores que adquiramos, están enraizados en nuestra herencia biológica y luego modificados mediante el aprendizaje. Al igual que con los principios anteriores, el refuerzo cultural está asentado en la evolución, en el desarrollo y en el aprendizaje. Los principios de la competencia y la cooperación culturales tienen raíces biológicas similares. Al haber evolucionado como animales sociales, los humanos están dotados de un equilibrio sutil entre impulsos competitivos y cooperativos. Nuestra naturaleza competitiva es el resultado de la necesidad de favorecer nuestra propia supervivencia y reproducción. Al competir por la comida, por la comodidad o por la atención, incrementaremos la probabilidad de sobrevivir y de dejar descendencia. La cooperación dentro de un grupo social también es importante, por lo que nos podemos beneficiar de la asistencia mutua: si cooperamos con nuestro compañero, veremos incrementadas las opciones de supervivencia y de prosperidad de nuestros hijos. La competencia y la cooperación interaccionan de todas las maneras posibles durante el cambio cultural, pero siempre estarán enraizadas en nuestro pasado biológico.
Lo mismo vale para el principio de la riqueza combinatoria: el cerebro está estructurado para operar de un modo combinatorio para integrar la información que llega de muchos impulsos. En el caso de los humanos, estos impulsos incluirían información procedente de otras personas, de lo que dicen y de lo que hacen. Como nuestras redes neurales se pasan información unas a otras, estos impulsos sociales refuerzan algunas conexiones neurales mientras debilitan a otras. Gracias a este proceso, el cerebro se modifica, lo que nos permite beneficiarnos de los logros de los demás y llegar a nuevas combinaciones en forma de lenguaje, utensilios o ideas. El inmenso espacio cultural por el que transita la especie humana está arraigado en el modo combinatorio con el que nuestro cerebro se desarrolla, funciona e interacciona.
Finalmente, el principio de la recurrencia también está asentado en nuestras características biológicas. Desde un punto de vista evolutivo, no es rentable estar satisfecho, sino que sería mejor buscar continuamente acciones que incrementen la probabilidad de supervivencia y reproducción. Como en muchos otros animales, nuestro cerebro está estructurado para buscar la mejor manera de actuar entre las opciones disponibles. Aunque mejoren las condiciones, la complacencia nunca conviene: se podrían desaprovechar oportunidades y correr el riesgo de que a otros les vaya mejor que a nosotros. En un conjunto determinado de opciones, tendemos a buscar la que concuerda mejor con nuestros valores: la mejor comida, la mejor pareja, el mejor utensilio, la mejor manera de comunicarse, el mejor modo de sortear problemas o la mejor explicación[138]. Las opciones disponibles y lo que valoramos pueden cambiar con el tiempo, pero no nuestra búsqueda de lo mejor. Incluso un monje de clausura puede buscar el mejor lugar para meditar o el mejor modo de vida. Una falta de interés por buscar lo mejor, sin preocuparse de lo que suceda, se considera una enfermedad, un signo de desconexión o de depresión.
Tal y como vimos con el aprendizaje DT (capítulo 7), aprendemos de una manera relacional, siempre ajustando nuestras expectativas al aprender de las discrepancias, por lo que a medida que algunas opciones o valores se fortalecen y afianzan, aparecen otros nuevos que conducen a nuevas rondas de refuerzo y competencia. Este desplazamiento continuo de las expectativas y de los valores es lo que mantiene la cultura en constante movimiento. Si alcanzásemos un punto en el que todo el mundo estuviera completamente satisfecho, entonces sería probable que se detuviera la cultura. Pero esto es muy poco probable porque tenemos una tendencia natural a estar cada vez más insatisfechos con algunos aspectos de la vida: nuestro cerebro busca continuamente discrepancias y las acciones que ayudarán a resolverlas. Además, si apareciese un reducto estancado de cultura humana, sería vulnerable a una invasión de un grupo competidor de personas menos satisfechas que buscan nuevos territorios o recursos. El principio de recurrencia, el contexto siempre cambiante del espacio cultural, está fundamentado en nuestra biología, en ese impulso continuo que nos lleva a encontrar la mejor manera de actuar entre las opciones disponibles.
Todos los ingredientes para el cambio cultural están fundamentados en nuestro pasado biológico y nos los encontraremos en otros animales sociales, desde las termitas a los perros. Pero con la evolución del Homo sapiens, los ingredientes se juntaron e interaccionaron con una fuerza particular. Nuestra capacidad para aprender, comunicarnos e interaccionar es mucho mayor que la de cualquier otro ser vivo. Un termitero constituye un esfuerzo colectivo magnífico, pero su forma básica solo cambiaría a escala evolutiva mediante las alteraciones en las reacciones instintivas de sus constructores. En cambio, nuestras casas y ciudades son muy diferentes de las moradas de nuestros ancestros, hace 10.000 años. No se debe a que haya cambiado algún gen, sino a las transformaciones culturales que surgen de nuestra capacidad para aprender, comunicarnos e interaccionar. A medida que avanza nuestro periplo cultural, nuestros progresos también han servido de realimentación para mejorar los diferentes ingredientes en los que se basan. El desarrollo de medios de comunicación y de transporte más eficaces, por ejemplo, impulsó la variabilidad de la población, la persistencia, el refuerzo, la competencia, la cooperación y la riqueza combinatoria, además de catalizar nuestro movimiento recurrente por el espacio cultural.
Hemos visto que la evolución, el desarrollo y el aprendizaje son tres ejemplos de la receta creativa para la vida que se enmarcan unos a otros. Los tres procesos tienen una forma parecida y también están conectados por la historia, de manera que el desarrollo se engasta en la evolución y el aprendizaje en ambos procesos. Ahora podemos ver el cambio cultural como un cuarto ejemplo de la receta enmarcado por sus predecesores. No solo se parece a las otras transformaciones al tener una receta creativa común, sino que está engastado en ellas.
Pero démosle una vuelta de tuerca más a la historia. Nuestros conocimientos sobre evolución, desarrollo y aprendizaje son producto de la cultura. Las teorías de Darwin o de Turing son resultado de nuestra herencia cultural durante siglos. Nuestro cuarto ejemplo de la receta es especial por algo muy concreto: la cultura humana no solo está enmarcada por los otros tres procesos, sino que gracias a ella conseguimos verlos a todos. Enmarca y está enmarcado por todos ellos. Sería como si Simbad, al contar su historia a los invitados, comenzase a leer en voz alta el libro Las mil y una noches que contiene su propia historia.
La cultura como sistema de encuadre no solo se aplica a la biología, sino a toda la ciencia. Las teorías de Newton y de Einstein son también un producto cultural, como las de Darwin o Turing. ¿Cómo podemos compaginar este doble aspecto de la cultura? ¿Qué debemos considerar que es más importante, el marco de la cultura para la ciencia, o el marco de la ciencia para la cultura? Para responder esta pregunta, quiero estudiar de dónde vienen algunas de nuestras ideas más básicas sobre el mundo.
§. Mundos posibles
Solemos decir que el espacio tiene tres dimensiones, que contiene objetos materiales con longitud, anchura y profundidad. Podemos imaginar objetos que tengan menos dimensiones, pero no tendrían una existencia material: un objeto bidimensional sería infinitamente delgado y, por lo tanto, insustancial. Pero no es tan fácil pensar en objetos que tengan más de tres dimensiones. No nos resulta fácil imaginar objetos cuatridimensionales[139]. ¿Esta limitación es una característica de nuestra perspectiva o del mundo físico que nos rodea?Miremos en primer lugar las diferentes maneras que usamos para evaluar los objetos. Cuando tenemos una manzana en la mano, podemos apreciar muchos de sus aspectos, como el color, el olor y la forma. Si se apaga la luz, dejaremos de verla, pero su aroma y la forma seguirían resultando evidentes a los sentidos del olfato y del tacto. Diríamos que la manzana todavía está ahí, pero que no podemos verla. De igual forma, si ya no pudiéramos oler la manzana por alguna razón, no diríamos que la manzana se esfumó porque seguiríamos viéndola y sintiéndola. Sin embargo, la situación es muy diferente si imaginamos que la manzana ya no resiste la prensión: si viéramos que los dedos la atraviesan cuando intentamos agarrarla, entonces concluiríamos que se habría convertido en un fantasma y que en realidad ya no existe. El modo en el que entramos en contacto con una manzana y la sentimos parece ser algo intrínseco a ella, mientras que los atributos como el aspecto visual y el olor son más circunstanciales.
Desde un punto de vista biológico, que atribuyamos más importancia a unos aspectos de los objetos que a otros está relacionado con lo que más importa para nuestra supervivencia y reproducción. Lo más importante para nosotros es el tipo de contactos que establecemos con los objetos que nos rodean. Comemos en contacto con la comida, nos reproducimos en contacto con otros humanos semejantes, y prolongamos nuestra vida al evitar el contacto con las cosas que podrían matarnos. El sentido del tacto nos proporciona el acceso más inmediato al contacto. Cuando tocamos algo, sentimos que es real porque el contacto nos causará un daño real o será bueno. Los sentidos como la vista son más indirectos, ya que no podemos comer ni reproducirnos con la vista; esta es solo importante en la medida en que nos ayuda a favorecer o evitar determinados contactos. La vista nos ayudará a entrar en contacto con la comida o con la pareja, y a evitarlo si se trata depredador. Tendemos a pensar que el tacto es el árbitro principal de la realidad, un juez de lo realmente importante, mientras la vista tiene una importancia secundaria. Si viviéramos como las plantas sería de otro modo, porque la exposición a la luz tendría consecuencias muy directas para la supervivencia, ya que dependeríamos de ella directamente para alimentarnos mediante la fotosíntesis; las cualidades visuales parecerían por tanto más fundamentales.
Algunos aspectos de la vista, como la forma de un objeto, están muy correlacionados con el tacto: podemos ver y sentir que una manzana es esférica. Estos aspectos compartidos contribuyen a nuestra visión tridimensional de los objetos. La vista y el tacto colaboran diciéndonos que no necesitamos más que tres dimensiones para describir la extensión de un objeto. Los aspectos visuales que no se correlacionan con el tacto no forman parte de esta descripción. El color de una manzana no es directamente relevante para lo que se siente, por lo que nuestras tres dimensiones son incoloras (y son inodoras y silenciosas por motivos parecidos).
Una buena manera de transmitir cuatro dimensiones es incorporar una cualidad adicional en el modo en el que experimentamos los contactos. Voy a utilizar el color como ejemplo. Empezaré por presentar el mundo del arcoíris. Se trata de un mundo como el nuestro, salvo que cada objeto tiene un único color (estrictamente hablando, un intervalo de color), que se encuentra en algún tramo del espectro visible. Algunos objetos son rojos, otros amarillos, etc. La peculiaridad más extraña del mundo del arcoíris es que los objetos solo chocan entre sí cuando tienen el mismo color. Por lo tanto, una persona roja puede chocarse con una mesa roja, pero puede atravesar una de color naranja. Mediante el tacto, la persona roja siente solo cosas rojas, aun cuando pueda ver objetos de otros colores. Esto ocurrirá tanto si la luz está encendida como apagada: una persona roja seguirá chocándose con una mesa roja en la oscuridad aunque no sea capaz de ver que era roja. Una característica más del mundo del arcoíris es que los humanos y otros animales pueden cambiar de color. Si una persona roja quiere dejar de tropezar con una mesa roja, pueden decidir que la rodeará o que la atravesará cambiando su propio color a naranja.
Si hubiésemos evolucionado en el mundo del arcoíris, podríamos pensar de forma natural que los objetos tienen cuatro dimensiones: las tres dimensiones espaciales y la cuarta del color. Estaríamos acostumbrados a los desplazamientos por el espacio y por los colores para encontrar comida y pareja. Para comer una manzana, no solo necesitaríamos movernos hacia ella, sino también cambiar nuestro color para que coincidiéramos, y si quisiésemos evitar a un depredador, tendríamos las opciones de cambiar de color o salir corriendo. Por supuesto, el depredador podría a su vez cambiar de color para coincidir con el nuestro, con lo que nos perseguiría en la dimensión del color igual que nos podría perseguir por el espacio. El cerebro sería un experto pensador cuatridimensional.
Podríamos pensar que el color actúa aquí más bien como el tiempo, al igual que algunas veces tratamos el tiempo como una dimensión más. Pero el tiempo es diferente porque no podemos alterarlo con nuestros propios esfuerzos y sentir las consecuencias. Por supuesto, podemos retroceder en el tiempo con los recuerdos, pero cuando lo hacemos, no chocamos con los objetos que recordamos: si sabemos que alguien se sentó en donde estamos sentados ahora, el recuerdo de ese suceso no nos hace chocar con dicha persona. En cambio, el mundo del arcoíris es genuinamente cuatridimensional porque podemos cambiar el color y sentir las consecuencias, tal y como podemos cambiar nuestra posición espacial para provocar o evitar un contacto.
Ejemplos como el mundo del arcoíris indican que la incapacidad para pensar en cuatro dimensiones no se debe a una limitación intrínseca del cerebro, sino al modo en el que interaccionamos con el mundo que nos rodea. Si nuestro mundo funcionase de acuerdo a las reglas del mundo del arcoíris, entonces habríamos evolucionado, desarrollado y aprendido a pensar en cuatro dimensiones en vez de en tres. Por supuesto, no vivimos en el mundo del arcoíris porque los objetos con colores diferentes no tienen la costumbre de atravesarse unos a otros. Vivimos en un mundo en el que nuestro patrón de contactos puede ser explicado con tres dimensiones. El origen de nuestro punto de vista no se encuentra simplemente en nuestro cerebro ni en el mundo que nos rodea, sino en el mundo con el que interaccionamos.
Vamos a llegar a la misma conclusión de otra manera. La teoría de la gravedad de Newton tiene algunas limitaciones. Como advirtió Einstein, la teoría comienza a fallar cuando los cuerpos se acercan a la velocidad de la luz y no consigue explicar correctamente el efecto de la gravedad sobre la propia luz[140]. Einstein trató estos problemas con sus teorías especial y general de la relatividad, que desafían algunas de nuestras suposiciones más preciadas sobre el espacio y el tiempo. Lo normal es que pensemos que el espacio y el tiempo son independientes entre sí, de manera que el tiempo pasa del mismo modo con independencia de lo rápido que nos movamos por el espacio. De acuerdo con la relatividad especial, el tiempo y el espacio están íntimamente conectados por lo que el tiempo transcurrirá de un modo distinto en los objetos que se están moviendo unos respecto a otros. Esto solo se consigue percibir cuando los objetos se acercan a la velocidad de la luz, por lo que en nuestra escala normal de existencia podemos ignorar dichos efectos relativistas. De igual forma, los efectos de la gravedad sobre la luz, como se describe mediante la relatividad general, se hacen más evidentes cuando las masas son muy grandes, como en un agujero negro donde la gravedad es tan fuerte que la luz no se puede escapar.
La relatividad y su concepto de espacio y tiempo nos resultan más abstractos y difíciles de entender que la teoría de Newton, lo que se debe a la manera en la que interaccionamos con el mundo y no a ninguna capacidad de abstracción inherente. Nuestro cerebro está organizado para hacer frente a los objetos que nos rodean, que no suelen acercarse a la velocidad de la luz cuando se mueven respecto a nosotros (la luz viaja a aproximadamente mil millones de kilómetros por hora) y, por lo tanto, los percibimos en una muy buena aproximación como si el espacio y el tiempo fueran independientes. Pero esto no funcionaría si trabajáramos de forma habitual a una escala diferente. Como escribe el astrónomo Martin Rees, «una inteligencia que pudiera transitar rápidamente por el universo —constreñida por las leyes físicas básicas, pero no por la tecnología actual— ampliaría sus intuiciones sobre el espacio y el tiempo para que incorporasen las consecuencias peculiares y aparentemente extravagantes de la relatividad»[141]. A esa inteligencia que habitaría en una escala muy superior a la nuestra, la teoría de la relatividad le resultaría más sencilla que nuestra percepción de independencia del espacio y del tiempo.
Necesitaremos una gimnasia imaginativa similar para el mundo de lo minúsculo. La materia está compuesta por muchísimos átomos. Solemos pensar que los átomos son como los objetos cotidianos, salvo que son extremadamente pequeños, como pelotas de ping-pong pequeñísimas. Sin embargo, esta visión fue derrocada por los descubrimientos de la mecánica cuántica: a escala atómica y de las partículas que la forman, como los electrones, ya no valen las suposiciones sobre los objetos cotidianos. No podemos, por ejemplo, saber exactamente dónde está un electrón si también queremos saber lo rápido que se mueve, simplemente porque resulta imposible detener un electrón en su camino y decir «aquí está». Cuanto más seguros estemos de la posición del electrón, más inseguros estaremos de lo rápido que se mueve. Comenzaremos a hablar en términos de probabilidad en vez de certeza, y la materia adquiere un aspecto entre difuso y borroso. La borrosidad disminuye a medida que se incrementa la masa del objeto, por lo que en el momento en que obtenemos objetos visibles, como las manzanas, compuestos por una cantidad ingente de átomos, el grado de incertidumbre se vuelve insignificante. Podemos apuntar a una manzana que reposa en un lugar concreto de una mesa porque el nivel de incertidumbre para algo con la masa de una manzana es tan diminuto que resulta imperceptible. Los efectos cuánticos siguen existiendo en la manzana, pero serán despreciables a menos que ampliemos la escala hasta la de los átomos que la forman. Al igual que nuestros conceptos de espacio y tiempo se desmontaban debido a los efectos relativistas cuando la velocidad o la masa se vuelven enormes, nuestro concepto habitual de los objetos y de la materia se desmorona por los efectos cuánticos cuando nos adentramos en la escala atómica.
Esto no significa que haya tres tipos fundamentales de realidad física, cada uno para las escalas pequeña, mediana y grande, sino que es una consecuencia del modo en el que normalmente interaccionamos con el mundo. Como humanos, adquirimos conceptos que nos permiten ocuparnos de los objetos que podemos ver y tocar con facilidad, como las manzanas o las montañas. Nuestro concepto de objetos con una posición definida en el espacio y en el tiempo es un marco de referencia extremadamente eficaz para hacer frente al mundo a esta escala. Sin embargo, los conceptos cotidianos empiezan a resultarnos menos apropiados cuando nos movemos a escalas muy alejadas de nuestra norma, tanto por grande como por pequeña. Para manejarnos con estas escalas, necesitaremos expandir nuestros conceptos de siempre, a menudo de un modo que nos resultará incómodo o desconcertante. Como nuestras ideas se basan en las interacciones con el mundo a una escala concreta, nos veremos forzados a pensar de un modo que parece más abstracto a medida que intentamos abarcar escalas más amplias.
Los ejemplos como el mundo del arcoíris, la relatividad y la mecánica cuántica resaltan lo que muchos filósofos ya habían apuntado[142]: no podemos acceder directamente a nuestro mundo, sino que siempre nos veremos forzados a mirarlo a través de marcos de referencia concretos que no son arbitrarios, sino que proceden del mundo y de nuestra interacción con él. La idea de espacio tridimensional tiene que ver con el modo en que interaccionamos con el mundo concreto en el que nos encontramos, de manera que si fuera diferente, también lo contemplaríamos de un modo distinto. Las teorías de la relatividad y de la mecánica cuántica también constituyen marcos de referencia para estudiar el mundo, y nos revelan que no es tan sencillo como nuestras interacciones cotidianas con él podrían hacernos pensar. No obstante, estos marcos de referencia se basan en nuestra interacción con el mundo, porque los científicos idearon estas teorías mediante la experimentación y la observación. No podemos separar con nitidez nuestra visión del mundo físico y el modo en el que interaccionamos con él.
Lo mismo valdrá para nuestras explicaciones científicas de la evolución, del desarrollo y del aprendizaje. No podemos describir estos procesos si no es mediante los marcos de referencia culturales que se basan en ellos. Esto no significa que nuestros puntos de vista científicos sean arbitrarios, sino que son el reflejo de la estructura del mundo y de nuestras interacciones con él. La relación entre nuestros cuatro ejemplos de la receta creativa para la vida no es una simple cadena unidireccional desde la ciencia a la cultura, o desde la cultura a la ciencia, sino que implica una interacción bidireccional entre nuestros puntos de vista y los procesos que los originaron.
§. Autorretrato de la naturaleza
Cézanne era un hombre obsesionado, y algunas de sus obsesiones eran más bien desafortunadas. Le aterrorizaba que lo tocaran, como comprobó Émile Bernard cuando un día cometió el error de intentar ayudarlo después de que se cayera[143]. Pero su obsesión con las manzanas es algo a lo que le estaremos eternamente agradecidos: pintó más de 30 bodegones con manzanas (véase la figura 83, lámina 14, por ejemplo). De acuerdo con el crítico Gustave Geffroy, Cézanne proclamó «Asombraré a París con una manzana»[144]. Su objetivo no era simplemente copiar manzanas, sino utilizar estos humildes objetos para explorar las relaciones con el color, la tonalidad y la forma. «Pintar la naturaleza», se le atribuye que dijo, «es dejar libre la esencia del modelo. La pintura no significa la copia sumisa de un objeto. El artista debe percibir y capturar la armonía existente en medio de otras muchas relaciones[145] ». Cada artista capturará estas armonías y relaciones a su modo. Basta con comparar la pintura de Cézanne con la manera en la que Renoir trató un objeto parecido (figura 84, lámina 15), hecha más o menos en la misma época. Las manzanas de Renoir parecen más suaves que las de Cézanne y carecen de su intensa calidad escultural, porque las vio y las pintó a su manera.
Figura 83. Bodegón con membrillo, manzanas y peras, Paul Cézanne, hacia 1885-1887. Véase la lámina 14.
Figura 84. Manzanas y peras, Pierre-Auguste Renoir, hacia 1885-1887. Véase la lámina 15.
Pero las conexiones también se establecen en la dirección opuesta, desde la manzana cultural hacia las otras. Nuestra cultura, que incluye nuestra perspectiva científica, proporciona el marco de referencia a través del cual vemos el mundo. Esto significa que no podemos fundamentar nuestra visión del mundo simplemente en la física, porque tan pronto como describimos el mundo físico, lo estaremos haciendo a través de un marco de referencia cultural concreto. Ni tampoco podemos cimentar nuestra visión del mundo meramente en la cultura, porque nuestros puntos de vista no flotan aislados, sino que están impregnados del modo en el que evolucionamos, nos desarrollamos y aprendemos. En vez de un argumento lineal con un comienzo simple, tenemos una interacción bidireccional cuyos marcos de referencia enmarcan y se enmarcan mutuamente. Puede parecer poco satisfactorio el que no seamos capaces de identificar un punto de partida absoluto, pero quizá se deba a la otra peculiaridad de nuestra herencia cultural: nos gusta que las historias tengan un comienzo, un nudo y un desenlace. Pero este no es el modo en que se estructura el mundo ni nuestro lugar dentro de él.
La interdependencia entre los marcos de referencia y lo que enmarcan no comenzó con los humanos: hemos de pensar que cada microorganismo constituye un marco de referencia que captura las relaciones con su entorno. El alga unicelular Chlamydomonas tiene proteínas Rubisco que encajan con la forma de las moléculas de dióxido de carbono, lo que les permite fijar carbono. También tiene flagelos de tipo látigo y una capacidad de respuesta que le permite nadar hacia la luz. Estas adaptaciones permiten que el microorganismo esté preparado para determinados aspectos de su mundo. Esta preparación se produjo gracias a una interacción bidireccional continua entre el organismo y el entorno. Los organismos enmarcan su entorno de maneras concretas, y el entorno entonces sirve de realimentación para seleccionar los marcos de referencia que se verán favorecidos por la selección natural. El periplo de las poblaciones por el espacio genético está íntimamente conectado con el entorno que las rodea. Gracias a este proceso, cada especie ha evolucionado con su propio y profuso grupo de peculiaridades, que constituye un modo peculiar de capturar las relaciones con su entorno, del mismo modo que cada artista tiene su propia perspectiva del mundo. Este emparejamiento entre el organismo y el entorno, entre el marco de referencia y lo enmarcado, surge gracias a la evolución, que fue nuestro primer ejemplo de la receta creativa para la vida.
Los otros marcos de referencia aparecen cuando la evolución da vida al desarrollo, la segunda versión de la receta creativa para la vida. A medida que los organismos pluricelulares evolucionaron, consiguieron capturar aspectos de su entorno cada vez más complejos. El sargazo vejigoso captura el movimiento del mar, la gravedad y el modo en el que incide la luz al crecer de una forma particular con distintos tipos celulares: ordena el mundo ordenándose a sí mismo. Los organismos capturan las relaciones espaciales del mismo modo que capturan los patrones temporales. La maleza florece según la duración del día, la hoja de la Venus atrapamoscas se cierra cuando un insecto deambula por ella, y una babosa de mar acaba por ignorar que se le está tocando continuamente. Los periplos de los embriones por el espacio del desarrollo, engendrado por la evolución, están conectados íntimamente con su entorno. Cada planta y animal pluricelulares proporcionan un retrato determinado del mundo que ha surgido mediante la evolución y el desarrollo.
Las respuestas al entorno se han vuelto particularmente elaboradas en los animales, y alteran constantemente lo que estos experimentan mediante sus propios movimientos. Muchos de estos organismos desarrollan un sistema nervioso y un cerebro que les permite aprender a través de la secuencia de acontecimientos que se van encontrando. Son capaces de predecir lo que es probable que les recompense o les castigue, y luego modificar su acción en consecuencia. Aprenden a calibrar sus acciones frente a sus efectos y a manejarse con más eficacia en el mundo que les rodea. Con la exploración y el sondeo del mundo, se anclan a sí mismos en él, enmarcándose al mismo tiempo a sí mismos y a su entorno. Como están continuamente resolviendo discrepancias y construyendo sobre las mismas, verán el mundo bajo una nueva perspectiva, y hallarán nuevos marcos de referencia que capturarán lo que les rodea. El aprendizaje representa otro modo de dividir el mundo a través de marcos de referencia neurales que se modifican según la experiencia.
La interdependencia entre el marco de referencia y lo enmarcado no es única de la cultura, sino que se encuentra en todos los casos de la receta creativa para la vida. La evolución, el desarrollo y el aprendizaje conducen a una organización de la materia, el organismo, que capta las relaciones con la materia de su entorno (incluidos otros organismos). Los organismos son materia que se enmarca a sí misma. La receta creativa para la vida proporciona los principios generales mediante los que surge tal auto enmarcación. Se trata de una receta para la auto descripción, una receta mediante la cual el mundo se retrata a sí mismo de distintas maneras. En la sociedad de los humanos, el autorretrato ha alcanzado un nuevo nivel: el modo en el que los organismos conciben su propio origen y su lugar en la naturaleza. Este autorretrato muestra algunas de nuestras propias peculiaridades como seres humanos. Pero el cuadro que aparece no es arbitrario, del mismo modo que un autorretrato de Rembrandt no es una colección arbitraria de pinceladas. Más bien la situación se parece a la litografía de M. C. Escher que muestra a un hombre que mira un cuadro del que forma parte (figura 85). Al igual que el hombre de Escher, nunca podemos salir de nuestro cuadro, pero esto no significa que no podamos contemplarlo e intentar comprender el mundo fascinante del cual somos una parte inseparable.

Figura 85. Galería de grabados, M. C. Escher, 1956.
En particular, quisiera agradecerle a mi colega Andrew Bangham su ánimo y los estimulantes debates desde las primeras etapas del libro. Sus conocimientos de programación junto con los de Andy Hanna también resultaron decisivos para desarrollar las ideas del reconocimiento y los programas de análisis de retratos utilizados en el capítulo 9. También estoy profundamente en deuda con Peter Dyan por ser generoso con su ayuda y consejos en los capítulos sobre el aprendizaje, y a mis padres, Doris y Ernesto Coen, por su apoyo infatigable y la lectura crítica de los muchos borradores que les he enviado.
Me gustaría agradecerle a Graeme Mitchison sus utilísimos comentarios y su ánimo, a Roger Carpenter su guía crítica sobre la ciencia de los movimientos oculares, a Chris Frith los comentarios útiles sobre la cognición, a Alicia Hidalgo su ayuda general y sus consejos sobre la neurobiología del desarrollo, a Jonathan Miller sus conversaciones estimulantes sobre el reconocimiento y el cambio cultural, y a Przemyslaw Prusinkiewicz las muchas conversaciones útiles y divertidas. Le estoy agradecido a Richard Kennaway por pasar a ordenador el Cézanne deformado y a Ellen Poliakoff por los datos sobre el seguimiento del ojo. Gracias también por sus útiles aportes a Katie Abley, Dennis Bray, Antonio Cuadrado-Fernández, Veronica Grieneisen, Martin Howard, Stan Marée, Marie Mirouze y David Stern. También me gustaría agradecerle a mi agente Peter Tallack su ayuda para dar forma al argumento del libro, a mi editor en PUP, Alison Kallet, su sabiduría y su capacidad para reencauzarme con tacto cuando fue necesario, y a la correctora Sheila Dean y a la editora Beth Clevenger por sus útiles comentarios. Por supuesto, me responsabilizo de cualquier error o interpretación errónea de este libro.
Gracias también a los miembros de mi laboratorio y a mi ayudante personal, Georgina van-Aswegen, por su apoyo durante el tiempo que estuve escribiendo el libro. Finalmente, no podría haber escrito este libro sin el apoyo continuo y la comprensión de mi esposa Lucinda y de mis hijos Pip, Timmy y Susie.-Acker, W. B. R. Some T’ang andPre-T’angTexts on Chinese Paintings. Dover, Nueva York, 1954.
-Allis, C. D. Epigenetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Nueva York, 2009.
-Bahrami, B., K. Olsen, et al. «Optimally Interacting Minds».Science 329 (5995): pp. 1081-1085, 2010.
-Ball, P. H2O: A Biography of Water. Phoenix Press, Londres, 2000.
-Balleine, B. W. y A. Dickinson. «Goal-Directed Instrumental Action: Contingency and Incentive Learning and Their Cortical Structures».Neuropharmacology 37, 1998, pp. 407-419.
-Bard, J. «A Unity Underlying the Different Zebra Striping Patterns». Journal of Zoology183, 1977, pp. 527-539.
-Barton, N. H., D. E. G. Briggs, et al. Evolution. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Nueva York, 2007.
-Bell, P. R., A. R. Hemsley. Green Plants: Their Origin and Diversity. Cambridge University Press, Cambridge, 1992.
-Berns, G. Satisfaction: The Science of Finding True Fulfillment. Henry Holt, Nueva York, 2005.
-Blackett, P. M. S. «Rutherford Memorial Lecture 1954».Physical Society Yearbook, 1955, pp. 13-22.
-Blackmore, S. The Meme Machine. Oxford University Press, Oxford, 1999.
-Bonner, J. T. Why Size Matters: From Bacteria to Blue Whales. Princeton University Press, Princeton, N. J., 2006.
-Boring, E. G. «A New Ambiguous Figure». American Journal of Psychology 42, 1930, pp. 444-445.
-Bray, D. Wetware: A Computer in Every Living Cell. Yale University Press, New Haven, Conn., 2009.
-Bridgeman, B., D. Hendry, et al. «Failure to Detect Displacement of the Visual World During Saccadic Eye Movements». Vision Research15, 1975, pp. 719-722.
-Brown, D. A. Leonardo da Vinci: Origins of a Genius. Yale University Press, New Haven, Conn., 1998.
-Bruce, V. y A. Young. In the Eye of the Beholder: The Science of Face Perception. Oxford University Press, Oxford, 2000.
-Carpenter, R. H. S. Movements of the Eyes. Pion, Londres, 1988.
—. Neurophysiology. Edward Arnold, Londres, 2003.
-Carroll, S. B. Endless Forms Most Beautiful: The New Science of Evo Devo and the Making of the Animal Kingdom. W. W. Norton, Nueva York, 2005.
-Carroll, S. B., J. K. Grenier, y S. Weatherbee. From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. Blackwell Science, Malden, Mass., 2001.
-Coen, E. The Art of Genes. Oxford University Press, Oxford, 1999 (hay trad. cast.: El arte de los genes, Biblioteca Budirán, Barcelona, 2007).
-Cook, M. y S. Mineka. «Observational Conditioning of Fear to Fear-Relevant -Versus Fear-Irrelevant Stimuli in Rhesus Monkeys». Journal of Abnormal Psychology 98 (4), 1989, pp. 448-459.
-Cootes, T. F., C. J. Taylor, et al. «Active Shape Models: Their Training and Application».Computer Vision and Image Understanding 61 (1), 1995, pp. 38-59.
-Corner, E. H. The Life of Plants. Weidenfeld and Nicolson, Londres, 1964.
-Coyne, J. A. «The Self-Centred Meme». Nature 398, 1999, pp. 767-768.
-Crick, F. «Neural Edelmanism».Trends in Neurosciences 12, 1989, pp. 240-248.
-Darwin, C. A Journal of Researches into the Natural History and Geology of the Countries Visited during the Voyage ofH. M. S. «Beagle» Round the World. Ward Lock, Londres, 1890.
—. The Autobiography of Charles Darwin and Selected Letters, ed. Francis Darwin. Dover, Nueva York, 1958.
—. The Descent of Man and Selection in Relation to Sex. John Murray, Londres, 19012.
-Daw, N. D., Y. Niv y P. Dayan. «Uncertainty-Based Competition Between Prefrontal and Dorsolateral Striatal Systems for Behavioral Control». Nature Neuroscience8 (12), 2005, pp. 1704-1711.
-Dawkins, R. The Selfish Gene. Oxford University Press, Oxford 1976 (hay trad. cast.:El gen egoísta, Salvat, Barcelona, 1993).
—. The Blind Watchmaker. Penguin, Londres y Nueva York, 1986 (hay trad. cast.: El relojero ciego, RBA coleccionables S.A., Barcelona, 2004).
-Dayan, P. y L. F. Abbott.Theoretical Neuroscience: Computational and Mathematical Modeling of Neural Systems. MIT Press, Cambridge, Mass., 2005.
-Dean, B. «An Explanatory Label for Helmets». Bulletin of the Metropolitan Museum of Art10, 1915, pp. 173-177.
—. «The Hobby of Collecting Ancient Armor». American Society of Arms Collectors Bulletin 70, 1994, pp. 24-28.
-Dennett, D. C. Darwin’s Dangerous Idea: Evolution and the Meanings of Life. Penguin, Londres, 1996.
-Diamantaras, K. I. y S. Y. Kung. Principal Component Neural Networks: Theory and Applications. Wiley, Nueva York, 1996.
-Diamond, J. Guns, Germs and Steel: A Short History of Everybody for the Last 13,000 Years . Vintage, Londres, 1998.
-Doran, M. Conversations with Cézanne. University of California Press, Berkeley, California, 2001.
-Eagleman, D. Incognito: The Secret Lives of the Brain. Cannongate, Edimburgo, 2011.
-Edelman, G. Bright Air, Brilliant Fire: On the Matter of Mind. Penguin, Londres, 1994.
-Eisenstein, E. L. The Printing Revolution in Early Modern Europe. Cambridge University Press, Cambridge, 1983.
-Eriksson, M. E. y A. J. Millar. «The Circadian Clock. A Plant’s Best Friend in a Spinning World».Plant Physiology 132, 2003, pp. 732-738.
-Etcoff, N. L., R. Freeman y K. R. Cave. «Can We Lose Memories of Faces? Content Specificity and Awareness in a Prosopagnosic». Journal of Cognitive Neuroscience 3, 1991, pp. 25-41.
-Euler, T., P. B. Detwiler y W. Denk. «Directionally Selective Calcium Signals in Dendrites of Starburst Amacrine Cells».Nature 418 (6900), 2002, pp. 845-852.
-Fitch, W. T. The Evolution of Language. Cambridge University Press, Cambridge, 2010.
-Forssell, D. C., ed. Perceptual Control Theory: Science and Applications —A Book of Readings . Living Control Systems Publishing: www.livingcontrolssystems.com, 2009.
-Foster, R. y L. Kreitzman. Seasons of Life: The Biological Rhythms That Enable Living Things to Thrive and Survive . Profile Books, Londres, 2010.
-Foster, R. G. y L. Kreitzman. Rhythms of Life: The Biological Clocks that Control the Daily Lives of Every Living Thing . Profile Books, Londres, 2005.
-Frith, C. Making up the Mind: How the Brain Creates our MentalWorld. Blackwell, Malden, Mass., 2007.
-Gallese, V., L. Fadiga, et al. «Action Recognition in the Premotor Cortex». Brain 119, 1996, pp. 593-609.
-Gavrilets, S. Fitness Landscapes and the Origin of Species. Princeton University Press, Princeton, N. J., 2004.
-Gayford, M. Man with a Blue Scarf: On Sitting for a Portrait by Lucian Freud. Thames and Hudson, Londres, 2010.
-Gladwell, M. Outliers: The Story of Success. Penguin, Londres y Nueva York, 2009.
-Gogolla, N., I. Galimberti y P. Coroni (2007). «Structural Plasticity of Axon Terminals in the Adult». Current Opinion in Neurobiology 17 (5), 2007, pp. 516-524.
-Gould, S. J. Ontogeny and Phylogeny. Harvard University Press, Cambridge Mass., 1977 (hay trad. cast.: Ontogenia y filogenia: la ley fundamental biogenética, Crítica, Barcelona, 2010).
—. Life’s Grandeur: The Spread of Excellence from Plato to Darwin. Jonathan Cape, Londres, 1996 (hay trad. cast.: La grandeza de la vida. La expansión de la excelencia de Platón a Darwin , Crítica, Barcelona, 1997).
-Green, A. A., J. R. Kennaway, et al. «Genetic Control of Organ Shape and Tissue Polarity».PLoS Biology 8(11), 2010, p. e1000537.
-Greene, B. The Fabric of the Cosmos. Alfred A. Knopf, Nueva York, 2004.
-Gregory, R. Mirrors in Mind. W. H. Freeman Spektrum, Oxford, 1997.
-Haldane, J. B. S. Possible Worlds and Other Essays. Chatto and Windus, Londres, 1927 (hay trad. cast.: Mundos posibles, José Janés, Barcelona, 1947).
-Hawkins, J. y S. Blakeslee. On Intelligence. Henry Holt, Nueva York, 2004.
-Haykin, S. Neural Networks and Learning Machines. Pearson Prentice Hall, Upper Saddle River, N. J., 2009.
-Hebb, D. O. The Organisation of Behaviour: a Neuropsychological Theory. Wiley, Nueva York, 1949 (hay trad. cast.: La organización de la conducta, Debate, Barcelona, 1985).
-Heidegger, M. Being and Time. Traducción al inglés de J. Macquarrie y E. Robinson. Basil Blackwell, Oxford, 1962 (hay trad. cast.:Ser y tiempo, Editorial Universitaria, Santiago de Chile, 2005 4).
-Helmholtz, H. Helmholtz’s Treatise on Physiological Optics. Vol. 3. Traducción al inglés J. P. C. Southall. Optical Society of America, Nueva York, 1924. Publicado por primera vez en 1866 como Handbuch der Physiologischen Optik.
-Hinton, G. E. y S. J. Nowlan. «How Learning Can Guide Evolution». Complex Systems1, 1987, pp. 495-502.
-Hodges, A. Alan Turing: The Enigma. Vintage, Londres, 1992.
-Howard, J. Darwin. Oxford University Press, Oxford, 1982.
-Hubbell, S. P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, Princeton, N. J., 2001.
-Hubel, D. H. y T. N. Wiesel. Brain and Visual Perception: The Story of a 25-YearCollaboration. Oxford University Press, Oxford, 2005.
-Irwin, D. E. «Information Integration Across Saccadic Eye Movements». Cognitive Psychology 23, 1991, pp. 420-456.
-Jones, F. M. «The Most Wonderful Plant in the World». Natural History 23 (6), 1923, pp. 589-596.
-Jones, J. The Lost Battles: Leonardo, Michelangelo and the Artistic Duel that Defined the Renaissance . Simon and Schuster, Londres, 2010.
-Juniper, B. E. y D. J. Mabberley.The Story of the Apple. Timber Press, Portland, Oregón, 2006.
-Juniper, B. E., R. J. Robins y D. M. Joel. The Carnivorous Plants. Academic Press, Londres, 1989.
-Kamin, L. J. «Predictability, Surprise, Attention and Conditioning». En Punishment and Aversive Behavior, ed. B. A. Campbell y R. M. Church. Appleton-Century-Crofts, Nueva York, 1969, pp. 279-296.
-Kandel, E. R. In Search of Memory: The Emergence of a New Science of Mind. W.W. Norton, Nueva York, 2006.
-Kant, I. Critique of Pure Reason, 1986. Traducción al inglés en 1986 por J. M. D. Meiklejohn. Dent, Londres y Melbourne. Publicada por primera vez en 1781 (hay trad. cast.: Crítica de la razón pura, Taurus, Madrid, 2005).
-Kemp, M. Leonardo. Oxford University Press, Oxford, 2004 (hay trad. cast: Leonardo, Fondo de Cultura Económica, México, 2006).
-Knoll, A. H. Life on a Young Planet: The First Three Billion Years of Evolution on Earth . Princeton University Press, Princeton, N. J, 2003.
-Koch, C. Biophysics of Computation. Oxford University Press, Nueva York, 1999.
Kondo, S. y T. Miura. «Reaction-Diffusion Model as a Framework for Understanding Biological Pattern Formation».Science 329 (5999), 2010, pp. 1616-1620.
-Konorski, J. Conditioned Reflexes and Neuron Organisation. Cambridge University Press, Cambridge, 1948.
-Kruse, K., M. Howard y W. Margolin. «An Experimentalist’s Guide to Computational Modelling of the Min System». Molecular Microbiology 63 (5), 2007, pp. 1279-1284.
-Lakoff, G. y M. Johnson.Metaphors We Live By. University of Chicago Press, Chicago, Illinois, 1980.
-Land, M. F. «Motion and Vision: Why Animals Move Their Eyes». Journal of Comparative Physiology A 185, 1999, pp. 341-352.
-Lane, N. Life Ascending: The Ten Great Inventions of Evolution. Profile, Londres, 2010.
-Lebrecht, D., M. Foehr, et al. «Bicoid Cooperative DNA Binding is Critical for Embryonic Patterning in Drosophila». Proceedings of the National Academy of Sciences of the United States of America 102 (37), 2005, pp. 13 176-13 181.
-Leroi, A. M. Mutants: On the Form, Varieties and Errors of the Human Body. Harper Collins, Londres, 2003.
-Levi-Montalcini, R. «Nerve Growth Factor 35 Years Later».Science 237, 2009, pp. 1154-1162.
-Li, N. y J. J. DiCarlo. «Unsupervised Natural Experience Rapidly Alters Invariant Object Representation in Visual Cortex». Science 321 (5895), 2008, pp. 1502-1507.
-Llinás, R. R. I of the Vortex. MIT Press, Cambridge, Mass., 2001.
MacCurdy, E. The Notebooks of Leonardo da Vinci. Reprint Society, Londres, 1938.
-MacGregor, N. A History of the World in 100 Objects. Allen Lane, Londres, 2010.
-Mayr, E. What Evolution Is. Phoenix Press, Londres, 2002.
-McNeill, J. R. y W. H. McNeill. The Human Web: A Bird’s-EyeView of World History. W.W. Norton, Nueva York y Londres, 2003.
-Meinhardt, H. The Algorithmic Beauty of Sea Shells. Springer-Verlag, Berlín, 1998.
-Miller, K. D., J. B. Keller y M. P. Stryker. «Ocular Dominance Column Development: Analysis and Simulation». Science 245, 1989, pp. 605-615.
-Milner, A. D. y M. A. Goodale.The Visual Brain in Action. Oxford University Press, Oxford, 1996.
-Montague, P. R., P. Dayan, y T. J. Sejnowski. «A Framework for Mesencephalic Dopamine Systems Based on Predictive Hebbian Learning». Journal of Neuroscience 16 (5), 1996, pp. 1936-1947.
-Montague, R. Your Brain Is (Almost) Perfect: How we Make Decisions. Plume, Nueva York, 2007.
-Morrison,V. The Art of George Stubbs. Grange Books, Londres, 1997.
-Mukamel, R., A. D. Ekstrom, et al. «Single-Neuron Responses in Humans during Execution and Observation of Actions».Current Biology 20, 2010, pp. 750-756.
-Murray, H. J. R. A History of Chess. Oxford University Press, Oxford, 1913.
Neisser, U. Cognition and Reality. W. H. Freeman, San Francisco, California, 1976.
-Nicholl, C. Leonardo da Vinci: The Flights of the Mind. Penguin, Londres, 2004.
Noble, D. The Music of Life: Biology beyond the Genome. Oxford University Press, Oxford, 2006.
-Nüsslein-Volhard, C. Coming to Life: How Genes Drive Development. Yale University Press, New Haven, Conn., 2006.
-Olshausen, B. A., C. H. Anderson y D. C. Van Essen. «A Neurobiological Model of Visual Attention and Invariant Pattern Recognition Based on Dynamic Routing of Information». Journal of Neuroscience 13 (11), 1993, pp. 4700-4719.
-Palmer, D. Origins: Human Evolution Revealed. Mitchell Beazley, Londres, 2010.
-Palubicki, W., K. Horel, et al. «Self-Organizing Tree Models for Image Synthesis». ACM Transactions on Graphics 28 (3), 2009, pp. 1-58.
-Pávlov, I. P. Conditioned Reflexes: An Investigation of the Physiological Activity of the Cerebral Cortex . Traducción y edición en inglés de G. V. Anrep. Oxford University Press, Oxford, 1927.
-Penfield, W. y T. Rasmussen. The Cerebral Cortex of Man. MacMillan, Nueva York, 1950.
-Pinker, S. The Language Instinct. Penguin, Londres, 1994.
-Prothero, R. E., ed. The Works of Lord Byron. Letters and Journals. Octagon Books, Nueva York, 1966.
-Ptashne, M. y A. Gann. Genes and Signals. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Nueva York, 2001.
-Purves, D., G. L. Augustine, et al. Neuroscience. Sinauer, Sunderland, Mass., 2008.
-Ramachnandran, V. S. The Tell-Tale Brain: Unlocking the Mystery of Human Nature. William Heinemann, Londres, 2011.
-Rapport, R. Nerve Endings: The Discovery of the Synapse. W. W. Norton, Nueva York, 2005.
-Rees, M. Just Six Numbers: The Deep Forces that Shape the Universe. Phoenix Press, Londres, 1999.
-Reichardt, W. «Autocorrelation, a Principle for the Evaluation of Sensory Information by the Central Nervous System». En Principles of Sensory Communication, W. A. Rosenblith ed. John Wiley, Nueva York, 1961, pp. 303-317.
-Reynolds, J. Discourses. Penguin, Londres, 1992. Publicado por primera vez en 1769.
-Ridley, M. The Red Queen: Sex and the Evolution of Human Nature. Viking, Londres, 1993.
—. The Rational Optimist: How Prosperity Evolves. Fourth Estate, Londres, 2010 (hay trad. cast.: El optimista racional, Taurus, Madrid, 2010).
-Robinson, A. Sudden Genius? The Gradual Path to Creative Breakthroughs. Oxford University Press, Oxford, 2010.
-Romo, R. y W. Schultz. «Dopamine Neurons of the Monkey Midbrain: Contingencies of Responses to Active Touch During Self-Initiated Arm Movements». Journal of Neurophysiology 63 (3), 1990, pp. 592-606.
-Rooke, J. E. y T. Xu. «Positive and Negative Signals Between Interacting Cells for Establishing Neural Fate».Bioessays 20 (3), 1998, pp. 209-214.
-Sachs, J. Lectures on the Physiology of Plants. Oxford University Press, Oxford, 1887.
Sacks, O. The Man Who Mistook his Wife for a Hat. Picador, Londres, 1986 (hay trad. cast.: El hombre que confundió a su mujer con un sombrero, Anagrama, Barcelona, 2004).
—. An Anthropologist on Mars. Picador, Londres, 1995.
-Sanes, D. H., T. A. Reh y W. A. Harris. Development of the Nervous System. Elsevier, Burlington, Mass., 2006.
-Schultz, W. (2010). «Dopamine Signals for Reward Value and Risk: Basic and Recent Data».Behavioral and Brain Functions 6: artículo 24, 2010, pp. 1-9 (en línea).
-Seugnet, L., P. Simpson, et al. «Transcriptional Regulation of Notch and Delta: Requirement for Neuroblast Segregation in Drosophila». Development 124 (10), 1997, pp. 2015-2025.
-Shenk, D. The Immortal Game: A History of Chess or How 32 Carved Pieces on a Board Illuminated Our Understanding of War, Art, Science, and the Human Brain . Souvenir Press, Londres, 2007.
-Sherrington, C. Man on His Nature. Cambridge University Press, Cambridge, 1951.
-Simons, P. The Action Plant. Blackwell, Oxford y Cambridge, 1992.
-Skinner, B. F. Science and Human Behavior. Macmillan, Nueva York, 1953.
Smith, J. M. y E. Szathmáry. The Major Transitions in Evolution. W. H. Freeman, Oxford, 1995.
-Smith, P. Interpreting Cézanne. Tate Publishing, Londres, 1996.
-Spirov, A., K. Fahmy, et al. «Formation of the Bicoid Morphogen Gradient: An RNA Gradient Dictates the Protein Gradient».Development 136 (4), 2009, pp. 605-614.
-Stern, D. Evolution, Development, and the Predictable Genome. Roberts, Greenwood Village, Colo., 2011.
-Stevens, J. K., R. C. Emerson, et al. «Paralysis of the Awake Human: Visual Perceptions».Vision Research 16, 1976, pp. 93-98.
-Stratton, G. M. «Vision without Inversion of the Retinal Image». Psychological Review 4, 1897, pp. 341-60.
-Straube, A., A. F. Fuchs, et al. «Characteristics of Saccadic Gain Adaptation in Rhesus Macaques». Journal of Neurophysiology 77, 1997, pp. 874-895.
-Stubbs, G. The Anatomy of the Horse. G. Heywood Hill, Londres, 1938. Originalmente impreso para el autor por J. Purser en 1766.
-Suri, R. E. y W. Schultz. «A Neural Network Model with Dopamine-Like Reinforcement Signal That Learns a Spatial Delayed Response Task». Neuroscience 91 (3), 1999, pp. 871-890.
-Sutton, R. «Learning to Predict by the Methods of Temporal Differences». Machine Learning 3, 1988, pp. 9-44.
-Sutton, R. S. y A. G. Barto. «Toward a Modern Theory of Adaptive Networks: Expectation and Prediction».Psychological Review 88, 1981, pp. 135-171.
-Swindale, N. V. «A Model for the Formation of Ocular Dominance Stripes». Proceedings of the Royal Society of London. Series B. 208, 1980, pp. 243-264.
-Toriyama, H. «The Behaviour of the Sensitive Plant in a Typhoon». Botanical Magazine79, 1966, pp. 427-428.
-Turing, A. «The Chemical Basis of Morphogenesis». Philosophical Transactions of the Royal Society of London. Series B. 237, 1952, pp. 37-72.
-Vetter, T. y T. Poggio. «Linear Object Classes and Image Synthesis From a Single Example Image». IEEE Transactions on Pattern Analysis and Machine Intelligence 19 (7), 1997, pp. 733-742.
-Von der Malsburg, C. «Development of Ocularity Domains and Growth Behaviour of Axon Terminals».Biological Cybernetics 32, 1979, pp. 49-62.
—. «Network Self-Organisation». En An Introduction to Neural and Electronic Networks, ed. S. F. Zornetzer, J. L. Davis y C. Lau. Academic Press, San Diego, California, 1990, pp. 421-432.
-Wallman, J. y A. Fuchs. «Saccadic Gain Modification: Visual Error Drives Motor Adaptation».Journal of Neurophysiology80, 1998, pp. 2405-2416.
-Wiskott, L. y T. J. Sejnowski. «Slow Feature Analysis: Unsupervised Learning of Invariances». Neural Computation 14, 2002, pp. 715-770.
-Wolpert, L. y C. Tickle. Principles of Development. Oxford University Press, Oxford, 2011.
-Zallen, J. A. «Planar Polarity and Tissue Morphogenesis». Cell129 (6), 2007, pp. 1051-1063.
-Zeki, S. A Vision of the Brain. Blackwell Scientific, Oxford, 1993.Figura 5/lámina 1. Retrato de Ambroise Vollard, 1899 (óleo sobre tela), de Paul Cézanne (1839-1906). Museo de la Ville de Paris, Museo del Petit Palais, París, Francia/Giraudon/The Bridgeman Art Library.
Figura 8. La creación de Adán, 1510, de Miguel Ángel Buonarroti. Capilla Sixtina, Vaticano, Roma.
Figura 9/lámina 3. La calle Montorgueil, París, Celebración del 30 de junio de 1878 (óleo sobre tela), 1878, de Claude Monet (1840-1926). Museo de Orsay, París, Francia/Giraudon/The Bridgeman Art Library.
Figura 10. La torre de Babel, 1563, de Pieter Brueghel el Viejo. Museo Kunsthistorisches, Viena.
Figura 11. Patrón obtenido al aplicar las reglas de Turing. Cortesía de Stan Marée.
Figura 24/lámina 4. El jardín de Les Lauves, 1906, de Paul Cézanne. La Colección Phillips, Washington DC.
Figura 26. Interacciones entre los temas y las variaciones en un embrión de la mosca del vinagre. Según Coen 1999, p. 300, figura 15.18.
Figura 27. Cebra, 1763, de George Stubbs (1724-1806). Centro Yale de arte británico, Colección de Paul Mellon.
Figura 28. Generación de las rayas en las diferentes especies de cebra. Carroll (2005), figura 9.8, 241. Cortesía de Sean Carroll.
Figura 29/lámina 5. Manzanas y galletas, hacia 1880, de Paul Cézanne. Museo Nacional de la Orangerie, París. Colección de Jean Walter y Paul Guillaume. Versión deformada por cortesía de Richard Kennaway.
Figura 30/lámina 6. Versión deformada de las Manzanas y galletas de Cézanne, por cortesía de Richard Kennaway.
Figura 32. Árbol generado por un programa de ordenador. De Palubicki, W., K. Horel, et al. (2009).
Figura 33. Simulación por ordenador de los cambios morfológicos de Dictyostelium. Cortesía de Stan Marée.
Figura 34. Anatomía del caballo, 1766, de George Stubbs. Decimotercera tabla anatómica. Real Academia de las Artes, Londres.
Figura 36. Corteza de cerebro en desarrollo. Según Sanes et al., 2006, p. 53.
Figura 37. Evolución de los yelmos. Según Dean 1915. Museo Metropolitano del Arte. Diagrama de Bashford Dean, Departamento de Armas y Armaduras. Tiene los derechos el Museo Metropolitano del Arte.
Figura 38. Sargazo vejigoso (Fucus vesiculosus). Según Bell, 1992, p. 92, figura 4.19.
Figura 39. Porción de césped, 1503, de Alberto Durero. Albertina, Viena.
Figura 41/lámina 7. El bibliotecario, 1565, de Giuseppe Arcimboldo. Colección de Eva Hökenberg, Estocolmo.
Figura 42/lámina 8. «Las cuatro estaciones», de Giuseppe Arcimboldo (1527-1593): Primavera, Verano, Otoño e Invierno, 1573 (óleo sobre tela). Louvre, París, Francia/Giraudon/The Bridgeman Art Library.
Figura 43. La hoja de la Venus atrapamoscas. De Juniper et al., 1989, p. 52.
Figura 44. Hojas de Mimosa pudica. De Sachs, 1887, p. 645, figura 373.
Figura 46. Dos neuronas. Según Rains, 2001, p. 32.
Figura 47. Neuronas en el ganglio abdominal de la babosa de mar Aplysia californica. Según Kandel, 2006, p. 224.
Figura 48. El cerebro de Aplysia. Según Kandel, 2006, p. 147.
Figura 50. El sistema nervioso humano. De Purves et al., 2008, p. 15, figura 1.1.
Figura 51. Áreas somatosensorial y motora. Según Penfield y Rasmussen, 1950.
Figura 52. Sibila de Libia, 1511, de Miguel Ángel Buonarroti. Capilla Sixtina, Vaticano, Roma.
Figura 57. Dibujo de bombardas en el folio 33.º del Códice Atlántico de Leonardo da Vinci, hacia 1504. Tienen los derechos la Veneranda Biblioteca Ambrosiana de Milán y la De Agostini Picture Library.
Figura 58/lámina 9. Bodegón con tetera, hacia 1869 (óleo sobre tela), de Paul Cézanne. Museo de Orsay, París, Francia. Crédito de la fotografía: Réunion des Musées Nationaux/Art Resource, NY. El seguimiento del ojo es cortesía de Ellen Poliakoff y Andrew Bangham.
Figura 62/lámina 2. Retrato de Ambroise Vollard (1868-1939), 1909 (óleo sobre tela), de Pablo Picasso (1881-1973). Museo Pushkin, Moscú, Rusia/Giraudon/The Bridgeman Art Library. Tienen los derechos en 2011 los herederos de Pablo Picasso/Artists Rights Society (ARS), Nueva York.
Figura 63. Mi mujer y mi suegra. De Boring, 1930.
Figura 70. Algunas áreas del cerebro implicadas en el procesamiento visual. Adaptado de Scientific American, nov. de 1999, p. 48.
Figura 72. Retrato de Maria Trip, 1639, de Rembrandt van Rijn, Rijksmuseum, Amsterdam. Retrato de una joven, 1917-1918, de Amedeo Modigliani. Colección privada.
Figura 75. Retrato de Jeanne Hébuterne, 1919, de Amedeo Modigliani. Art & Fragances S.A. Cham/Suiza. Oscar Miestchaninoff, 1917, de Amedeo Modigliani. Colección privada.
Figura 78. Patrones de dominancia ocular en la corteza visual primaria del mono macaco. De D. H. Hubel y T. N. Wiesel, Conferencia Ferrier. Proc. R. Soc. Lond. B 198, 1977, pp. 1-59, figura 24(a).
Figura 79/lámina 11. San Jerónimo y el león en el monasterio, 15011509 (óleo sobre tela), de Vittore Carpaccio (hacia 1460/5-1523/6). Escuela de San Giorgio degli Schiavoni, Venecia, Italia/Giraudon/The Bridgeman Art Library.
Figura 80/lámina 12. Hércules y la Hidra, hacia 1475 (pintura al temple sobre madera), de Antonio Pollaiuolo (1432/3-1498). Galería de los Uffizi, Florencia, Italia/Alinari/The Bridgeman Art Library.
Figura 81/lámina 13. Tobías y el ángel, 1470-1480, de Andrea del Verrocchio (1436-1488) y Leonardo da Vinci (1452-1519). National Gallery, Londres. Tienen los derechos National Gallery, Londres/Art Resource, NY. Tobías y el ángel (pintura al temple sobre madera), de Antonio Pollaiuolo y Piero Pollaiuolo. Galería Sabauda, Turín, Italia/Alinari/The Bridgeman Art Library.
Figura 83/lámina 14. Bodegón con membrillo, manzanas y peras, hacia 1885-1887 (óleo sobre tela), de Paul Cézanne (1839-1906). Legado de Charles A. Loeser, 1952. Asociación histórica de la Casa Blanca (Colección de la Casa Blanca): 523.
Figura 84/lámina 15. Manzanas y peras, hacia 1885-1887, de Pierre-Auguste Renoir (1841-1919). Museo de la Orangerie, París. Colección de Jean Walter y Paul Guillaume. Crédito de la fotografía: Réunion des Musées Nationaux/Art Resource, NY.
Figura 85. Galería de grabados, 1956, de M. C. Escher. Tiene los derechos The M. C. Escher Company-Holland. Todos los derechos reservados. www.mcescher.com.

LÁMINA 1. Retrato de Ambroise Vollard, Paul Cézanne, 1899.

LÁMINA 2. Retrato de Ambroise Vollard, Pablo Picasso, 1909.

LÁMINA 3. La calle Montorgueil, París, celebración del 30 de junio de 1878, Claude Monet, 1878.

LÁMINA 4. El jardín de Les Lauves, Paul Cézanne, 1906.

LÁMINA 5. Manzanas y galletas, Paul Cézanne, hacia 1880, y una versión deformada ocasionada por el mayor crecimiento de las regiones amarillas.

LÁMINA 6. La versión deformada de Manzanas y galletas de Cézanne cuando se deja que el lienzo se combe en tres dimensiones.

LÁMINA 7. El bibliotecario, Giuseppe Arcimboldo, 1565.

LÁMINA 8. «Las cuatro estaciones»: Primavera (arriba a la izquierda), Verano (arriba a la derecha), Otoño(abajo a la izquierda) e Invierno (abajo a la derecha), Giuseppe Arcimboldo, 1573.

LÁMINA 9. Movimientos oculares registrados durante veinte segundos mientras se observa Bodegón con tetera, de Paul Cézanne, hacia 1869.

LÁMINA 10. Dos de las tendencias capturadas por las neuronas de correlaciones de la colección de 179 retratos (véanse las pp. 265-267).

LÁMINA 11. San Jerónimo y el león en el monasterio, Vittore Carpaccio, 1501-1509.

LÁMINA 12. Hércules y la Hidra, Antonio Pollaiuolo, hacia 1475.


LÁMINA 13. Arriba: Tobías y el ángel, Andrea del Verrochio y Leonardo da Vinci, 1470-1480. Abajo: Tobías y el ángel, Antonio Pollaiuolo y Piero Pollaiuolo, 1460.
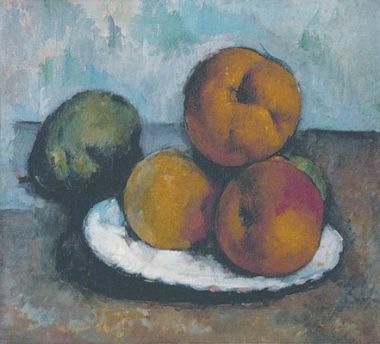
LÁMINA 14. Bodegón con membrillo, manzanas y peras, Paul Cézanne, hacia 1885-1887.

LÁMINA 15. Manzanas y peras, Pierre-Auguste Renoir, hacia 1885-1887
Notas: Dado el amplio alcance de este libro, solo es posible proporcionar un conjunto selecto de referencias. He diferenciado entre las referencias a las que podrá acceder cualquier lector general y la bibliografía técnica más especializada.